You might also like
- Essentials of Marketing Research 5th Edition Zikmund Test BankDocument18 pagesEssentials of Marketing Research 5th Edition Zikmund Test Bankbrittanylamsfwkpbmzce100% (18)
- Clinical Psychology Notes No. 1Document2 pagesClinical Psychology Notes No. 1Jerine Bonus ApostolNo ratings yet
- Carroll Essays Biochem 2013 Autophagy Ageing ChapterDocument15 pagesCarroll Essays Biochem 2013 Autophagy Ageing ChapterSKNo ratings yet
- Autophagy and Disease: Always Two Sides To A Problem: Invited ReviewDocument19 pagesAutophagy and Disease: Always Two Sides To A Problem: Invited ReviewCLPHtheoryNo ratings yet
- Nutrient Regulation of MTORC1 at A Glance JCS 2019Document6 pagesNutrient Regulation of MTORC1 at A Glance JCS 2019Osvaldo VillarNo ratings yet
- The Regulation of Aging - Does Autophagy Underlie Longevity - PMCDocument15 pagesThe Regulation of Aging - Does Autophagy Underlie Longevity - PMCParth RanaNo ratings yet
- ApoptosisDocument3 pagesApoptosisMega KapitanNo ratings yet
- Diet, Autophagy, and Cancer A ReviewDocument15 pagesDiet, Autophagy, and Cancer A ReviewAhmad Romdhoni Bin SalamNo ratings yet
- Metastasis and MAPK Pathways: Molecular SciencesDocument35 pagesMetastasis and MAPK Pathways: Molecular SciencesTehmeena BegumNo ratings yet
- Targeted Protein Degradation: Mechanisms, Strategies and ApplicationDocument13 pagesTargeted Protein Degradation: Mechanisms, Strategies and ApplicationJulián Tibaqué SepúlvedaNo ratings yet
- Mechanisms of Action of AntibioticsDocument4 pagesMechanisms of Action of AntibioticsClarissa Marie LimmangNo ratings yet
- Free Naa DESaM - Naa PrajaluDocument14 pagesFree Naa DESaM - Naa PrajaluAmarendar DeviniNo ratings yet
- Targeting Mitochondria For Cancer TherapyDocument18 pagesTargeting Mitochondria For Cancer Therapysatya_chagantiNo ratings yet
- ART Action and Resistance Mechanisms of Antibiotics A Guide For CliniciansDocument16 pagesART Action and Resistance Mechanisms of Antibiotics A Guide For CliniciansHECTORIBZAN ACERO SANDOVALNo ratings yet
- Misfolded ProteinsDocument8 pagesMisfolded ProteinsAfaq AhmadNo ratings yet
- Fibroblasts Modulate The Tumor Cell Motility and Their mTOR/S6K1 Phosphorylation Status in VitroDocument10 pagesFibroblasts Modulate The Tumor Cell Motility and Their mTOR/S6K1 Phosphorylation Status in VitroАнна ШаповаловаNo ratings yet
- Crosstalk Mechanism Between Degradative Pathways and Death PathwaysDocument27 pagesCrosstalk Mechanism Between Degradative Pathways and Death PathwaysKrupasindhu DindaNo ratings yet
- Confounding Factors From Inducible Systems For Spatiotemporal Gene Expression RegulationDocument4 pagesConfounding Factors From Inducible Systems For Spatiotemporal Gene Expression RegulationJessica Asitimbay ZuritaNo ratings yet
- FEBS Letters - 2020 - Eastman - The Palette of Techniques For Cell Cycle AnalysisDocument15 pagesFEBS Letters - 2020 - Eastman - The Palette of Techniques For Cell Cycle Analysissigmafranco8No ratings yet
- The Role of STAT3 in AutophagyDocument12 pagesThe Role of STAT3 in AutophagyeNo ratings yet
- Autoimmunity and AutophagyDocument18 pagesAutoimmunity and AutophagybuttercupsashNo ratings yet
- Shaping The Regulation of The p53 mRNA Tumour Suppressor: The Co-Evolution of Genetic SignaturesDocument16 pagesShaping The Regulation of The p53 mRNA Tumour Suppressor: The Co-Evolution of Genetic SignaturesSRIBDE STANLEYNo ratings yet
- Barbosa MC Et Al 2018Document21 pagesBarbosa MC Et Al 2018maria carolina barbosaNo ratings yet
- Oxidative Phosporylation in Cancer CellsDocument9 pagesOxidative Phosporylation in Cancer CellsM Naufal IlmiNo ratings yet
- Autophagy and Intermittent Fasting The Connection For Cancer TherapyDocument7 pagesAutophagy and Intermittent Fasting The Connection For Cancer TherapyYoBjjNo ratings yet
- Activation of Senescence in Critically Ill Patients: Mechanisms, Consequences and Therapeutic OpportunitiesDocument10 pagesActivation of Senescence in Critically Ill Patients: Mechanisms, Consequences and Therapeutic OpportunitiesGdfgdFdfdfNo ratings yet
- Review Article Autophagy in Acute Pancreatitis: Organelle Interaction and Microrna RegulationDocument12 pagesReview Article Autophagy in Acute Pancreatitis: Organelle Interaction and Microrna RegulationCésar CarvalloNo ratings yet
- Autophagy-Independent FunctionsDocument18 pagesAutophagy-Independent Functionselibb346No ratings yet
- Molecular Chaperones: Advanced Article R. John EllisDocument7 pagesMolecular Chaperones: Advanced Article R. John EllisazzaassNo ratings yet
- Resistensi Dan Cara Kerja AntiboticsDocument6 pagesResistensi Dan Cara Kerja AntiboticsChristiadi karlin12No ratings yet
- Properties and Functions ofDocument18 pagesProperties and Functions ofNivetha NagarajanNo ratings yet
- Uloga Autofagije U Imunskom Odgovoru I Autoimunskim BolestimaDocument7 pagesUloga Autofagije U Imunskom Odgovoru I Autoimunskim BolestimaHeather LewisNo ratings yet
- Chapter+2Cell+Proliferation Onco CeliaDocument34 pagesChapter+2Cell+Proliferation Onco CeliaPaula C GuimaraesNo ratings yet
- Wolfbam 2Document4 pagesWolfbam 2Michael WolfsonNo ratings yet
- Plasma Membrane Integrity: Implications For Health and DiseaseDocument29 pagesPlasma Membrane Integrity: Implications For Health and DiseaseJose Manuel Diaz MolinaNo ratings yet
- Autophagy Cellular and Molecular MechanismsDocument10 pagesAutophagy Cellular and Molecular MechanismsWilliam LeeNo ratings yet
- Ijms 16 17193Document38 pagesIjms 16 17193Afaq AhmadNo ratings yet
- 12 - Autophagy in Human DiseasesDocument13 pages12 - Autophagy in Human DiseasesAndres Felipe Hurtado BautistaNo ratings yet
- ArtigoDocument12 pagesArtigoGabrielNo ratings yet
- 10 1016@j Molmed 2019 10 009Document18 pages10 1016@j Molmed 2019 10 009Elena AmaralNo ratings yet
- Mitochondrial Respiratory ComplexesDocument30 pagesMitochondrial Respiratory ComplexesDouglas DíazNo ratings yet
- Protac 2Document14 pagesProtac 2muruganvvkNo ratings yet
- MTOR Is A Key Modulator of Ageing and Age-Related Disease: Nature January 2013Document9 pagesMTOR Is A Key Modulator of Ageing and Age-Related Disease: Nature January 2013SBNo ratings yet
- CMSA Medical PG Entrance Coaching Institute: CM SADocument13 pagesCMSA Medical PG Entrance Coaching Institute: CM SAAmarjeet KaurNo ratings yet
- News & Views: Surviving Import FailureDocument2 pagesNews & Views: Surviving Import FailureatharvatanksaleNo ratings yet
- Action and Resistance Mechanisms of Antibiotics - A Guide For CliniciansDocument6 pagesAction and Resistance Mechanisms of Antibiotics - A Guide For CliniciansJonatas SilvaNo ratings yet
- Immunometabolism in The Single-Cell Era: PerspectiveDocument16 pagesImmunometabolism in The Single-Cell Era: PerspectiveyasbiloNo ratings yet
- Bio305 Molecular Biology Summary 08024665051Document35 pagesBio305 Molecular Biology Summary 08024665051solomon obasogieNo ratings yet
- Menezes Et Al 2013Document20 pagesMenezes Et Al 2013Carina MenezesNo ratings yet
- 1 s2.0 S0962892414000865 MainDocument9 pages1 s2.0 S0962892414000865 Mainpfrancesco.soaveNo ratings yet
- Proteins and Cell Regulation Vol 03 - RHO Family GTPases, 1E (2005)Document306 pagesProteins and Cell Regulation Vol 03 - RHO Family GTPases, 1E (2005)DiahaNo ratings yet
- Journal of Biomedical Science: Molecular Mechanisms of Cell Proliferation Induced by Low Power Laser IrradiationDocument16 pagesJournal of Biomedical Science: Molecular Mechanisms of Cell Proliferation Induced by Low Power Laser IrradiationMohamed ElshounyNo ratings yet
- Immunotoxins and Cancer Therapy: Zheng Li, Tao Yu, Ping Zhao and Jie MaDocument7 pagesImmunotoxins and Cancer Therapy: Zheng Li, Tao Yu, Ping Zhao and Jie Mayota_ahlyNo ratings yet
- tmpC8C9 TMPDocument6 pagestmpC8C9 TMPFrontiersNo ratings yet
- Autophagy and Mitophagy in Cellular Damage ControlDocument5 pagesAutophagy and Mitophagy in Cellular Damage Controljiazhuo liNo ratings yet
- Molecular Mechanisms of Autophagy and Implications in Liver DiseaseDocument15 pagesMolecular Mechanisms of Autophagy and Implications in Liver DiseaseWaldo IzyNo ratings yet
- Role and Mechanisms of Mitophagy in Liver DiseasesDocument30 pagesRole and Mechanisms of Mitophagy in Liver DiseasescarolinaNo ratings yet
- Action and Resistance Mechanisms of Antibiotics: A Guide For CliniciansDocument6 pagesAction and Resistance Mechanisms of Antibiotics: A Guide For CliniciansAlejandra SánchezNo ratings yet
- Mitochondrial ApoptosisDocument5 pagesMitochondrial ApoptosisRuheen MariamNo ratings yet
- Minsung Kim@manchester Ac UkDocument1 pageMinsung Kim@manchester Ac UkAtif NaeemNo ratings yet
- Mitochondrial Membrane PermeabilizationDocument66 pagesMitochondrial Membrane PermeabilizationGus Tavo BrugesNo ratings yet
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsFrom EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneNo ratings yet
- SHANSEP Approach For Slope Stability Assessments of River Dikes in The NetherlandsDocument9 pagesSHANSEP Approach For Slope Stability Assessments of River Dikes in The NetherlandsKen LiewNo ratings yet
- CopyrightsDocument31 pagesCopyrightsxomilaaNo ratings yet
- Foundations of Economics Test ItemsDocument43 pagesFoundations of Economics Test ItemsFelix Tagud AraraoNo ratings yet
- InvitationDocument6 pagesInvitationAmaria MurilloNo ratings yet
- Self-Study Worksheet III Isomerism ANSWERSDocument2 pagesSelf-Study Worksheet III Isomerism ANSWERSkjjkimkmkNo ratings yet
- SOP For Filling Liquid Cylinder Using Pump Transfer Method Rev 1 PDFDocument9 pagesSOP For Filling Liquid Cylinder Using Pump Transfer Method Rev 1 PDFYASH ENGINEERING SOLUTIONSNo ratings yet
- 27th Nov2020 Daily Global Regioanl Local Rice E-Newsltter (Un-Edited Version)Document90 pages27th Nov2020 Daily Global Regioanl Local Rice E-Newsltter (Un-Edited Version)Mujahid AliNo ratings yet
- Towards SPM Practice Paper 1: 1. Given One of The Roots of The Quadratic Equation 3. A Quadratic Function Is Given AsDocument2 pagesTowards SPM Practice Paper 1: 1. Given One of The Roots of The Quadratic Equation 3. A Quadratic Function Is Given Aszamriu8No ratings yet
- Untitled DocumentDocument4 pagesUntitled Documentallyssa abrogenaNo ratings yet
- Nikos Salingaros-Complexity and Urban Coherence PDFDocument35 pagesNikos Salingaros-Complexity and Urban Coherence PDFIacubenco AlexandrNo ratings yet
- Acid-And Acid/Heat Coagulated Cheese: J A Lucey, University of Wisconsin-Madison, Madison, WI, USADocument8 pagesAcid-And Acid/Heat Coagulated Cheese: J A Lucey, University of Wisconsin-Madison, Madison, WI, USAمیلاد قاسمیNo ratings yet
- Unit: 5 Meaning of Accounting StandardsDocument13 pagesUnit: 5 Meaning of Accounting Standardsrajapriya 41No ratings yet
- Đề Ôn Thi Vào 10 2Document7 pagesĐề Ôn Thi Vào 10 2Quỳnh PhạmNo ratings yet
- HDFC Software Requirement Specification SrsDocument24 pagesHDFC Software Requirement Specification SrsAshish Roushan100% (1)
- Unit 3 Controlling The ConversationDocument2 pagesUnit 3 Controlling The ConversationdefiaaaNo ratings yet
- 3 Properties of Real Numbers ChartDocument1 page3 Properties of Real Numbers ChartwlochNo ratings yet
- Single-Phase ARD Series Elevator Automatic Rescue DeviceDocument11 pagesSingle-Phase ARD Series Elevator Automatic Rescue Deviceaheneke1No ratings yet
- Help Seeking As A Self-Regulated Learning Strategy: January 2013Document14 pagesHelp Seeking As A Self-Regulated Learning Strategy: January 2013Agus PratomoNo ratings yet
- Defoamer Test Method: 1 Ross-Miles Foam TesterDocument2 pagesDefoamer Test Method: 1 Ross-Miles Foam Testeradnan aslamNo ratings yet
- MSG.05.Equations and InequalitiesDocument7 pagesMSG.05.Equations and InequalitiesNicola NichelleNo ratings yet
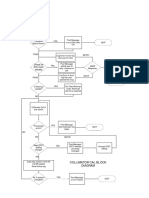
- Check For Dirty Mylar Window: Collimator Cal Block DiagramDocument1 pageCheck For Dirty Mylar Window: Collimator Cal Block DiagramMai Thanh SơnNo ratings yet
- OpenStax - Psychology - CH15 PSYCHOLOGICAL DISORDERSDocument42 pagesOpenStax - Psychology - CH15 PSYCHOLOGICAL DISORDERSAngelaNo ratings yet
- Basic Interior Design RulesDocument25 pagesBasic Interior Design RulesMadalina-Irina Tanaselea100% (2)
- Leadership and Followership NoteDocument11 pagesLeadership and Followership NoteDzihlornu PreciousNo ratings yet
- Procedure SDI TestDocument2 pagesProcedure SDI TestChairul ImamNo ratings yet
- Wise, Bolls, & Schaefer, 2008Document17 pagesWise, Bolls, & Schaefer, 2008Robert DumitruNo ratings yet
- 2CDC446001D0201 Busbar SystemsDocument36 pages2CDC446001D0201 Busbar SystemsBorislav ChavdarovNo ratings yet
- Safe Flight Aoa Quick ReferenceDocument4 pagesSafe Flight Aoa Quick ReferenceGary MedinaNo ratings yet