You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5813)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Statistics for Bioinformatics: Methods for Multiple Sequence AlignmentFrom EverandStatistics for Bioinformatics: Methods for Multiple Sequence AlignmentNo ratings yet
- An Introduction to Biotechnology: The Science, Technology and Medical ApplicationsFrom EverandAn Introduction to Biotechnology: The Science, Technology and Medical ApplicationsRating: 4.5 out of 5 stars4.5/5 (2)
- Tetrahedron Reports on Organic Chemistry: Volume 1.1-10From EverandTetrahedron Reports on Organic Chemistry: Volume 1.1-10Derek BartonNo ratings yet
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967From EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterNo ratings yet
- Cholinergic Ligand InteractionsFrom EverandCholinergic Ligand InteractionsD. J. TriggleNo ratings yet
- Coordination Chemistry—XVI: XVIth International Conference on Coordination ChemistryFrom EverandCoordination Chemistry—XVI: XVIth International Conference on Coordination ChemistryNo ratings yet
- Molecular Diseases: FEBS Federation of European Biochemical Societies: 12th Meeting, Dresden, 1978From EverandMolecular Diseases: FEBS Federation of European Biochemical Societies: 12th Meeting, Dresden, 1978G. JacobaschNo ratings yet
- Biochemical and Pharmacological Roles of Adenosylmethionine and the Central Nervous System: Proceedings of an International Round Table on Adenosylmethionine and the Central Nervous System, Naples, Italy, May 1978From EverandBiochemical and Pharmacological Roles of Adenosylmethionine and the Central Nervous System: Proceedings of an International Round Table on Adenosylmethionine and the Central Nervous System, Naples, Italy, May 1978Vincenzo ZappiaNo ratings yet
- Mass Spectrometry of Natural Products: Plenary Lectures Presented at the International Mass Spectrometry Symposium on Natural Products, Rehovot, Israel, 28 August - 2 September 1977From EverandMass Spectrometry of Natural Products: Plenary Lectures Presented at the International Mass Spectrometry Symposium on Natural Products, Rehovot, Israel, 28 August - 2 September 1977B. SklarzNo ratings yet
- PAKs, RAC/CDC42 (p21)-activated Kinases: Towards the Cure of Cancer and Other PAK-dependent DiseasesFrom EverandPAKs, RAC/CDC42 (p21)-activated Kinases: Towards the Cure of Cancer and Other PAK-dependent DiseasesHiroshi MarutaNo ratings yet
- Mechanisms in Radiobiology: Multicellular OrganismsFrom EverandMechanisms in Radiobiology: Multicellular OrganismsMaurice ErreraNo ratings yet
- Cell Biology Assays: ProteinsFrom EverandCell Biology Assays: ProteinsFanny JaulinNo ratings yet
- Handbook of GlycomicsFrom EverandHandbook of GlycomicsRichard D. CummingsNo ratings yet
- Azepines, Part 1From EverandAzepines, Part 1Andre RosowskyNo ratings yet
- Organic Synthesis - 2: Plenary Lectures Presented at the Second International Symposium on Organic SynthesisFrom EverandOrganic Synthesis - 2: Plenary Lectures Presented at the Second International Symposium on Organic SynthesisS. SarelNo ratings yet
- Progress in Biophysics and Molecular BiologyFrom EverandProgress in Biophysics and Molecular BiologyJ. A. V. ButlerNo ratings yet
- Applied Biophysics for Drug DiscoveryFrom EverandApplied Biophysics for Drug DiscoveryDonald HuddlerNo ratings yet
- Interpretation of the Ultraviolet Spectra of Natural Products: International Series of Monographs on Organic ChemistryFrom EverandInterpretation of the Ultraviolet Spectra of Natural Products: International Series of Monographs on Organic ChemistryNo ratings yet
- Carbon-13 NMR Spectroscopy of Biological SystemsFrom EverandCarbon-13 NMR Spectroscopy of Biological SystemsNicolau BeckmannNo ratings yet
- Helicases from All Domains of LifeFrom EverandHelicases from All Domains of LifeRenu TutejaNo ratings yet
- Sulfurtransferases: Essential Enzymes for LifeFrom EverandSulfurtransferases: Essential Enzymes for LifeNoriyuki NagaharaNo ratings yet
- Reproduction in Domestic AnimalsFrom EverandReproduction in Domestic AnimalsH. H. ColeNo ratings yet
- Natural Products Analysis: Instrumentation, Methods, and ApplicationsFrom EverandNatural Products Analysis: Instrumentation, Methods, and ApplicationsVladimir HavlicekNo ratings yet
- Ecdysone: From Metabolism to Regulation of Gene ExpressionFrom EverandEcdysone: From Metabolism to Regulation of Gene ExpressionMary BownesNo ratings yet
- Regulation of Macromolecular Synthesis By Low Molecular Weight MediatorsFrom EverandRegulation of Macromolecular Synthesis By Low Molecular Weight MediatorsGebhard KochNo ratings yet
- From Globular Proteins to AmyloidsFrom EverandFrom Globular Proteins to AmyloidsIrena Roterman-KoniecznaNo ratings yet
- Neuroendocrine Perspectives: Volume 3From EverandNeuroendocrine Perspectives: Volume 3Eugenio E. MüllerNo ratings yet
- Mathematical Biology: A Conference on Theoretical Aspects of Molecular ScienceFrom EverandMathematical Biology: A Conference on Theoretical Aspects of Molecular ScienceNo ratings yet
- Live Cell Assays: From Research to Regulatory ApplicationsFrom EverandLive Cell Assays: From Research to Regulatory ApplicationsNo ratings yet
- Fluctuations and Non-Linear Wave Interactions in Plasmas: International Series in Natural PhilosophyFrom EverandFluctuations and Non-Linear Wave Interactions in Plasmas: International Series in Natural PhilosophyNo ratings yet
- The Use Of Radioautography in Investigating Protein SynthesisFrom EverandThe Use Of Radioautography in Investigating Protein SynthesisC LeblondNo ratings yet
- Metabolic Inhibitors V1: A Comprehensive TreatiseFrom EverandMetabolic Inhibitors V1: A Comprehensive TreatiseR HochsterNo ratings yet
- Formation and Fate of Cell OrganellesFrom EverandFormation and Fate of Cell OrganellesKatherine Brehme WarrenNo ratings yet
- Valor Nutrimental de Los Productos de Origen AnimalDocument8 pagesValor Nutrimental de Los Productos de Origen Animalmaleny sanchezNo ratings yet
- Risque de La Biologie MoléculaireDocument35 pagesRisque de La Biologie Moléculaireمولاي أمحمد عكرميNo ratings yet
- Informe de BioquimicaDocument32 pagesInforme de BioquimicaJarys Daniela De La RosaNo ratings yet
- Trabajo-Arbol Filogenetico de La VidaDocument20 pagesTrabajo-Arbol Filogenetico de La VidaEdwin CondoriNo ratings yet
- Proyecto Sisistemas de Gestion Ambiental SabadoDocument52 pagesProyecto Sisistemas de Gestion Ambiental SabadoMargarita Cardnas OrozcoNo ratings yet
- Ficha de Indagación - AdnDocument3 pagesFicha de Indagación - AdnJimena Renata Urbina LoayzaNo ratings yet
- Practica 3 de ProteinasDocument10 pagesPractica 3 de ProteinasMichelle Andrea Villamar MotaNo ratings yet
- BIOGERONTOLOGIA (1) (Autoguardado)Document14 pagesBIOGERONTOLOGIA (1) (Autoguardado)Magnolia Yenny Mamani CcunoNo ratings yet
- Pre NutrientesDocument43 pagesPre NutrientesDaniela Beatríz SosaNo ratings yet
- Post Lab 2 BioquiDocument6 pagesPost Lab 2 BioquiPablo GonzalezNo ratings yet
- Alimentación Material OctubreDocument105 pagesAlimentación Material OctubreAnddres Polo100% (1)
- Alimenti e Ritmi OrmonaliDocument58 pagesAlimenti e Ritmi Ormonalipaolo contini100% (1)
- Compuestos Nitrogenados No ProteicosDocument2 pagesCompuestos Nitrogenados No ProteicosPrinci P Real50% (2)
- Valoracion de La Calidad PL1Document12 pagesValoracion de La Calidad PL1Jairon Terres QuispeNo ratings yet
- Tratamento Nutricional em Crianças Menores de 2 Anos Com Alergia Á Proeína Do Leite de VacaDocument33 pagesTratamento Nutricional em Crianças Menores de 2 Anos Com Alergia Á Proeína Do Leite de Vacajuliana silvaNo ratings yet
- Taller PDFDocument5 pagesTaller PDFKeyti De MojicaNo ratings yet
- Alimentación Durante El Primer Año de VidaDocument18 pagesAlimentación Durante El Primer Año de VidaDIANA JOCELYN HUALLPA CONTRERASNo ratings yet
- Que Es VidaDocument4 pagesQue Es VidaKIMBERLY GUADALUPE CARMONA RODRIGUEZNo ratings yet
- Genética BacterianaDocument4 pagesGenética BacterianaAndrea GarciaNo ratings yet
- Acetilcisteina Mec AçDocument4 pagesAcetilcisteina Mec AçCamilla CristinaNo ratings yet
- Desintoxicacion KitchariDocument32 pagesDesintoxicacion KitchariDenisseAgcNo ratings yet
- Problemset PDFDocument69 pagesProblemset PDFTrương ThiênNo ratings yet
- Taller ProteínasDocument5 pagesTaller ProteínasDiego RodriguezNo ratings yet
- Clase 1 Segundo CincuentaDocument78 pagesClase 1 Segundo CincuentaJonathanNo ratings yet
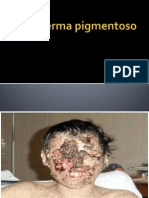
- Xeroderma PigmentosoDocument18 pagesXeroderma Pigmentosojosse_rodrigues_silvNo ratings yet
- Alimento Balanceados Cerdos La MolinaDocument9 pagesAlimento Balanceados Cerdos La MolinaBeatriz FalNo ratings yet
- Ração Humana PDFDocument7 pagesRação Humana PDFFaby MaiaNo ratings yet
- Informe Naturaleza Quimica ProtoplasmaDocument14 pagesInforme Naturaleza Quimica ProtoplasmaKevin AjilaNo ratings yet
- Derivados MelDocument34 pagesDerivados MelPaula Soares de MedeirosNo ratings yet
- Wykład MIĘSODocument88 pagesWykład MIĘSOela57635No ratings yet