You might also like
- Max Lab ReportDocument1 pageMax Lab ReportKallu PrasadNo ratings yet
- Global Status Report On Blood Safety and Availability 2021Document184 pagesGlobal Status Report On Blood Safety and Availability 2021Jessa MaeNo ratings yet
- Proceedings of the First International Congress of Parasitology: Roma, 21-26 September 1964From EverandProceedings of the First International Congress of Parasitology: Roma, 21-26 September 1964Augusto CorradettiNo ratings yet
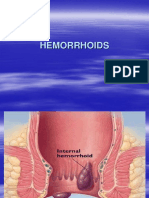
- 24 Anorectal ConditionsDocument55 pages24 Anorectal ConditionsRaisa CleizeraNo ratings yet
- A Crystallin Fold in The Interleukin - MeyerDocument17 pagesA Crystallin Fold in The Interleukin - Meyerana.pinho.souzaNo ratings yet
- Obata BasosDocument9 pagesObata BasosAynurNo ratings yet
- Department of Bio-Informatics 3 Semester (Morning) : Subject ImmunologyDocument10 pagesDepartment of Bio-Informatics 3 Semester (Morning) : Subject ImmunologyShaher Bano MirzaNo ratings yet
- A Mouse Model of Ovalbumin Allergen Accumulation of Peribronchial Mast Cells inDocument9 pagesA Mouse Model of Ovalbumin Allergen Accumulation of Peribronchial Mast Cells inEduardo MendesNo ratings yet
- Molecular Prevalence of Anaplasma Phagocytophilum in Sheep From IraqDocument8 pagesMolecular Prevalence of Anaplasma Phagocytophilum in Sheep From IraqZoleida BastidasNo ratings yet
- EosinófilosDocument13 pagesEosinófilosdkbritobNo ratings yet
- Familial Outbreak of Yersinia Enterocolitica Serotype O9 Biotype 2Document3 pagesFamilial Outbreak of Yersinia Enterocolitica Serotype O9 Biotype 2Diego TulcanNo ratings yet
- Eosinophils Protect Nematode Parasite Larvae by Regulating Local ImmunityDocument20 pagesEosinophils Protect Nematode Parasite Larvae by Regulating Local ImmunityLeonardo DonatiNo ratings yet
- Aina Velez (Pseudo-Research Proposal) PFT in Gen Bio 2Document7 pagesAina Velez (Pseudo-Research Proposal) PFT in Gen Bio 2Aina VelezNo ratings yet
- Adler, Et Al. 1999. Candidate Vaccine Antigens and Genes in P MultocidaDocument8 pagesAdler, Et Al. 1999. Candidate Vaccine Antigens and Genes in P MultocidakrodriguezNo ratings yet
- Cells Expression in Hematopoietic Progenitor E2F4 Modulates Differentiation and GeneDocument10 pagesCells Expression in Hematopoietic Progenitor E2F4 Modulates Differentiation and GenerinkuNo ratings yet
- Poirel 2018Document27 pagesPoirel 2018Leidy GarciaNo ratings yet
- 1 s2.0 S0002944010648523 MainDocument8 pages1 s2.0 S0002944010648523 MaindpcamposhNo ratings yet
- Occurrence of Gram-Negative Bacteria in Hens EggsDocument8 pagesOccurrence of Gram-Negative Bacteria in Hens EggsmeitaNo ratings yet
- Feline Infectious Peritonitis ABCD Guidelines On PDocument12 pagesFeline Infectious Peritonitis ABCD Guidelines On Pxn5q8nckvkNo ratings yet
- Siracusa Et Al. 2013. RevisionDocument15 pagesSiracusa Et Al. 2013. RevisionMelissa Rivera EscotoNo ratings yet
- Mora 2006Document8 pagesMora 2006kevin gelaudeNo ratings yet
- International Journal For Parasitology: Musa A. Hassan, Aude-Anais Olijnik, Eva-Maria Frickel, Jeroen P. SaeijDocument8 pagesInternational Journal For Parasitology: Musa A. Hassan, Aude-Anais Olijnik, Eva-Maria Frickel, Jeroen P. SaeijwindaNo ratings yet
- Regilating BCLCDocument11 pagesRegilating BCLCAnna GozaliNo ratings yet
- J Immunol 2007 Rydström 5789 801Document14 pagesJ Immunol 2007 Rydström 5789 801susana1616No ratings yet
- Enzootic Abortion in Ewes: A Review of Recent Developments in DiagnosticsDocument3 pagesEnzootic Abortion in Ewes: A Review of Recent Developments in DiagnosticsErnesto VersailleNo ratings yet
- Isolation of Actinobacillus Pleuropneumoniae From Pigs in IsraelDocument6 pagesIsolation of Actinobacillus Pleuropneumoniae From Pigs in IsraelDylan Warmfuzzies SalesNo ratings yet
- Nadif CurrOpinAllergyClin Immunol 2013 PDFDocument22 pagesNadif CurrOpinAllergyClin Immunol 2013 PDFMustafa AmmarNo ratings yet
- Basophils and Allergic Inflammation: Mechanisms of Allergic DiseasesDocument13 pagesBasophils and Allergic Inflammation: Mechanisms of Allergic DiseasesBasideu ByinajuNo ratings yet
- Loss of The Interleukin-6 Receptor Causes Immunodeficiency, Atopy, and Abnormal Inflammatory ResponsesDocument13 pagesLoss of The Interleukin-6 Receptor Causes Immunodeficiency, Atopy, and Abnormal Inflammatory ResponsesSyed Irfan RazaNo ratings yet
- 09 J Immunol 2010 Silverman 5011 22 LeishmaniaDocument13 pages09 J Immunol 2010 Silverman 5011 22 LeishmanialilianapradaNo ratings yet
- 1 s2.0 S0740002016304555 Main PDFDocument5 pages1 s2.0 S0740002016304555 Main PDFDIEGO FERNANDO TULCAN SILVANo ratings yet
- Differences Between IL-4R Deficiente Mice Reveal A Role For IL-13 in The Regulation of Th2 ResponsesDocument4 pagesDifferences Between IL-4R Deficiente Mice Reveal A Role For IL-13 in The Regulation of Th2 ResponsesIanca AndradeNo ratings yet
- REVIEWA Naplasma Marginale and Anaplasma Phagocytophilum-Rickettsiales Pathogens of Veterinary and Publichealth SignificanceDocument17 pagesREVIEWA Naplasma Marginale and Anaplasma Phagocytophilum-Rickettsiales Pathogens of Veterinary and Publichealth Significanceyudhi arjentiniaNo ratings yet
- 2018 Brucella Canis and Public Health RiskDocument6 pages2018 Brucella Canis and Public Health RiskAleida RomeroNo ratings yet
- Through A TLR5-and Flagellin Induces Antibody Responses: J ImmunolDocument11 pagesThrough A TLR5-and Flagellin Induces Antibody Responses: J ImmunolSEBASTIAN JOSE VEGA MARCELONo ratings yet
- Lin, Z. (2018) - A Metalloprotease Homolog Venom Protein From A Parasitoid Wasp Suppresses The Toll Pathway in Host HemocytesDocument15 pagesLin, Z. (2018) - A Metalloprotease Homolog Venom Protein From A Parasitoid Wasp Suppresses The Toll Pathway in Host HemocytesJanina PuenteNo ratings yet
- Salmonella Yoruba 1-s2.0-S0001706X22003126-mainDocument14 pagesSalmonella Yoruba 1-s2.0-S0001706X22003126-mainAYODELE ADESOJINo ratings yet
- Gamal Abdelgaber Mohammed Younis:: Sara Abdallah Mohammed FarhoudDocument6 pagesGamal Abdelgaber Mohammed Younis:: Sara Abdallah Mohammed FarhoudMohammed AbdallahNo ratings yet
- Jimmunol 1202354Document15 pagesJimmunol 1202354everton.monteiroNo ratings yet
- Staphylococcus Aureus Downregulates IP-10 Production and Prevents Th1 Cell Recruitment 2017Document11 pagesStaphylococcus Aureus Downregulates IP-10 Production and Prevents Th1 Cell Recruitment 2017oscarbio2009No ratings yet
- LWT - Food Science and Technology: Anna Zadernowska, Wioleta Chaje Cka-WierzchowskaDocument5 pagesLWT - Food Science and Technology: Anna Zadernowska, Wioleta Chaje Cka-WierzchowskaDiego TulcanNo ratings yet
- Proofs EuJMI 03 258Document10 pagesProofs EuJMI 03 258rizgah jawasNo ratings yet
- AnaplasmaDocument10 pagesAnaplasmabryanpes71No ratings yet
- Allergology International: Kensuke Miyake, Hajime KarasuyamaDocument10 pagesAllergology International: Kensuke Miyake, Hajime KarasuyamaponbohacopNo ratings yet
- Saprolegnia Diclina IIIA and S. Parasitica Employ Different Infection Strategies When Colonizing Eggs of Atlantic Salmon, Salmo Salar LDocument10 pagesSaprolegnia Diclina IIIA and S. Parasitica Employ Different Infection Strategies When Colonizing Eggs of Atlantic Salmon, Salmo Salar LcamilaNo ratings yet
- Neuropatologi Keracunan Organofosfat Pada Sapi: S Dan IDocument12 pagesNeuropatologi Keracunan Organofosfat Pada Sapi: S Dan IloviNo ratings yet
- Sabino R. 2016. Antifungal Susceptibility of 175 Aspergillus Isolates From Various Clinical and Enviromental SourcesDocument17 pagesSabino R. 2016. Antifungal Susceptibility of 175 Aspergillus Isolates From Various Clinical and Enviromental SourcesPaulette DlNo ratings yet
- Auto0711 1335Document13 pagesAuto0711 1335chocosherNo ratings yet
- Candida Albicans Iff11, A Secreted Protein Required For Cell WallDocument7 pagesCandida Albicans Iff11, A Secreted Protein Required For Cell Wall_meiantiNo ratings yet
- Eosinophils in Immunity and Disease Roles and Regulation of GastrointestinalDocument8 pagesEosinophils in Immunity and Disease Roles and Regulation of GastrointestinalBasideu ByinajuNo ratings yet
- Articulo 7 Mig OkDocument11 pagesArticulo 7 Mig Okoscarbio20090% (1)
- Salmonella in Laying HenDocument10 pagesSalmonella in Laying HenDaranee Nak-opatNo ratings yet
- Survey of Scid Gene and Immunological Status in Arabian Mares and FoalsDocument10 pagesSurvey of Scid Gene and Immunological Status in Arabian Mares and FoalsIJAR JOURNALNo ratings yet
- Garb Ett 2012Document8 pagesGarb Ett 2012Amir HayatNo ratings yet
- Isolation of Fungal Species and Aflatoxin Detection in Fermented ProductsDocument4 pagesIsolation of Fungal Species and Aflatoxin Detection in Fermented ProductsdodecadidoNo ratings yet
- PDF KVFD 1168Document6 pagesPDF KVFD 1168DindaNo ratings yet
- Intoxicación Por VoriconazolDocument2 pagesIntoxicación Por VoriconazolMariana fotos FotosNo ratings yet
- 14th ICI Abstract BookDocument4 pages14th ICI Abstract BookMuhammadGagasSasongkoNo ratings yet
- 9-M2 Polarization of Murine Peritoneal Macrophages (2016)Document9 pages9-M2 Polarization of Murine Peritoneal Macrophages (2016)Hadjer RezigNo ratings yet
- Identification of A Staphylococcal Complement Inhibitor With Broad Host Specificity in Equid Staphylococcus Aureus StrainsDocument10 pagesIdentification of A Staphylococcal Complement Inhibitor With Broad Host Specificity in Equid Staphylococcus Aureus StrainsChlo14No ratings yet
- Articulo 6 Ip-10okDocument11 pagesArticulo 6 Ip-10okoscarbio2009No ratings yet
- 3.03.05 - Avian - MycoDocument16 pages3.03.05 - Avian - MycoMehdi HassanNo ratings yet
- Sarcopenia Facts and Figures 2Document5 pagesSarcopenia Facts and Figures 2davidtuanandartuaNo ratings yet
- Classifying Giant Cell Lesions A Review.21Document5 pagesClassifying Giant Cell Lesions A Review.21shehla khanNo ratings yet
- Cir 0000000000000757Document458 pagesCir 0000000000000757Nona Suci rahayuNo ratings yet
- Eyes That Look Down and Out A Case Series On Isolated Third Nerve PalsyDocument4 pagesEyes That Look Down and Out A Case Series On Isolated Third Nerve PalsyIJAR JOURNALNo ratings yet
- BSN Psy. Version CDocument28 pagesBSN Psy. Version CImraan KhanNo ratings yet
- 2pdfnclex ReviewDocument187 pages2pdfnclex Reviewgarvita100% (1)
- Lesson 1 Reviewer in PmlsDocument10 pagesLesson 1 Reviewer in PmlsCharisa Joyce AgbonNo ratings yet
- AdsfDocument7 pagesAdsfNarito NianNo ratings yet
- "The Color of Health" by Common Ground HealthDocument27 pages"The Color of Health" by Common Ground HealthPatrick CrozierNo ratings yet
- JH CERILLES STATE COLLEGE - Post TestDocument7 pagesJH CERILLES STATE COLLEGE - Post TestCrystal Ann TadiamonNo ratings yet
- SCORES and CRITERIAS For Last Minute by DR RRMDocument52 pagesSCORES and CRITERIAS For Last Minute by DR RRMShweta Achuthan KuttyNo ratings yet
- Urinary Tract Infections (Utis) in Children Are Fairly Common, But Not Usually Serious. They Can Be Effectively Treated With AntibioticsDocument2 pagesUrinary Tract Infections (Utis) in Children Are Fairly Common, But Not Usually Serious. They Can Be Effectively Treated With AntibioticsWiljohn de la CruzNo ratings yet
- .Format of Note Making For Class 11 CBSE With ExampleDocument6 pages.Format of Note Making For Class 11 CBSE With ExampleRonak kadam100% (2)
- MS 1 Mastery TestDocument16 pagesMS 1 Mastery TestRika MaeNo ratings yet
- IJCRR Nov2020 Vol12 Issue21Document7 pagesIJCRR Nov2020 Vol12 Issue21Aravind KumarNo ratings yet
- Introduction To PharmacologyDocument30 pagesIntroduction To PharmacologyCandy crush100% (1)
- Reye SyndromeDocument10 pagesReye SyndromeDanil KhairulNo ratings yet
- Comparative Assessment of Conventional Periodontal Probes and CejDocument44 pagesComparative Assessment of Conventional Periodontal Probes and CejAnushree NingombamNo ratings yet
- Equine Field SurgeryDocument6 pagesEquine Field SurgeryRuchiNo ratings yet
- Arise RadioDocument15 pagesArise RadioRajanNo ratings yet
- Research Secondary HypertensionDocument51 pagesResearch Secondary HypertensionDomeng TalksNo ratings yet
- 4 6030444038488853198 PDFDocument110 pages4 6030444038488853198 PDFSyeda Seri RazaNo ratings yet
- Case ReportDocument9 pagesCase ReportJonatan DelgadoNo ratings yet
- New Guidelines For The Diagnosis of Paediatric Coeliac DiseaseDocument3 pagesNew Guidelines For The Diagnosis of Paediatric Coeliac DiseaseDiana NicaNo ratings yet
- Botox Course ManualDocument178 pagesBotox Course ManualMaged AbbasNo ratings yet
- Discover 12.2020Document68 pagesDiscover 12.2020nguyennamxmNo ratings yet
- E003335 Full PDFDocument3 pagesE003335 Full PDFOmer FarooqNo ratings yet