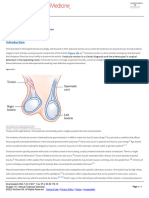
You might also like
- Balancing PediatricDocument13 pagesBalancing PediatricIsmiNo ratings yet
- Invasive Enterobacter Sakazakii Disease in Infants: Anna B. Bowen and Christopher R. BradenDocument5 pagesInvasive Enterobacter Sakazakii Disease in Infants: Anna B. Bowen and Christopher R. BradenRickyNo ratings yet
- Risk Factors For Hypoxic Ischemic Encephalopathy in ChildrenDocument5 pagesRisk Factors For Hypoxic Ischemic Encephalopathy in ChildrenSaqib AtherNo ratings yet
- 10 1056@NEJMoa1907423Document11 pages10 1056@NEJMoa1907423paredesol.modulo2.unipolNo ratings yet
- 2017 Article 141 Part2Document5 pages2017 Article 141 Part2Karina MonasaNo ratings yet
- Nejm - Eritropoyetina para Neuroprotección en PrematurosDocument11 pagesNejm - Eritropoyetina para Neuroprotección en PrematurosRonald MoralesNo ratings yet
- Antibiotics 10 00580Document11 pagesAntibiotics 10 00580Thu An Ngô ThịNo ratings yet
- Risk Factors For Necrotizing Enterocolitis Associated MortalityDocument7 pagesRisk Factors For Necrotizing Enterocolitis Associated MortalityvaleriaNo ratings yet
- Novel Interferon-Gamma Assays For Diagnosing Tuberculosis in Young Children in IndiaDocument10 pagesNovel Interferon-Gamma Assays For Diagnosing Tuberculosis in Young Children in IndiaRennyRayNo ratings yet
- 456 FullDocument4 pages456 FullDino GoncalvesNo ratings yet
- NIH Public Access: Author ManuscriptDocument8 pagesNIH Public Access: Author ManuscriptJuliet TyeNo ratings yet
- Summaries of Evidence in Support of Proposed Recommendations FinalDocument25 pagesSummaries of Evidence in Support of Proposed Recommendations FinalodyNo ratings yet
- A. Brief Resume of Intended WorkDocument6 pagesA. Brief Resume of Intended WorkRabi DhakalNo ratings yet
- Immune Responses To Measles and Mumps Vaccination of Infants at 6, 9, and 12 MonthsDocument10 pagesImmune Responses To Measles and Mumps Vaccination of Infants at 6, 9, and 12 MonthsManiru Aliyu GiganeNo ratings yet
- The NeoUpdates - DecDocument7 pagesThe NeoUpdates - DecDr Satish MishraNo ratings yet
- Nqy 012Document6 pagesNqy 012Pablo MoraNo ratings yet
- International Journal of Developmental NeuroscienceDocument6 pagesInternational Journal of Developmental NeuroscienceFarin MauliaNo ratings yet
- Adult Hippocampal Neurogenesis Is Regulated by The Mi - 2015 - Biological PsychiDocument3 pagesAdult Hippocampal Neurogenesis Is Regulated by The Mi - 2015 - Biological PsychiNoor AliNo ratings yet
- Early-Onset Sepsis: A Cohort Study Comparing Serial Clinical Observation With Laboratory TestingDocument7 pagesEarly-Onset Sepsis: A Cohort Study Comparing Serial Clinical Observation With Laboratory Testingcarmengmz22No ratings yet
- Solati 2015Document25 pagesSolati 2015Arthur Rocha GomesNo ratings yet
- Sensitivity of Histological Chorioaminionitis and Premature Rupture of Membranes For Neonatal Sepsis and Its Risk FactorsDocument6 pagesSensitivity of Histological Chorioaminionitis and Premature Rupture of Membranes For Neonatal Sepsis and Its Risk Factorsizza istqmhNo ratings yet
- The Effect of Probiotics in Prevention of Necrotising Enterocolitis in Preterm Neonates in Comparison With Control GroupDocument4 pagesThe Effect of Probiotics in Prevention of Necrotising Enterocolitis in Preterm Neonates in Comparison With Control GroupchachaNo ratings yet
- T-Helper (1) Cytokines Increase During Early Pregnancy in Women With A History of Recurrent Spontaneous AbortionDocument5 pagesT-Helper (1) Cytokines Increase During Early Pregnancy in Women With A History of Recurrent Spontaneous AbortionChu Chu gaNo ratings yet
- Faecal Calprotectin (FC) in Babies Born To Oher With or Without IBDocument2 pagesFaecal Calprotectin (FC) in Babies Born To Oher With or Without IBMartinaNo ratings yet
- Estacion EsDocument12 pagesEstacion EsLuis Quito CaroNo ratings yet
- Normal Values For Mature and Immature in Very Preterm BabiesDocument3 pagesNormal Values For Mature and Immature in Very Preterm BabiesSukma EffendyNo ratings yet
- Hiv 2023Document10 pagesHiv 2023Neonatología HGO4No ratings yet
- Role of Dexamethasone in Neonatal Meningitis 2012Document6 pagesRole of Dexamethasone in Neonatal Meningitis 2012jose alberto gamboaNo ratings yet
- Articulos Sepsis Neonatal TempranaDocument29 pagesArticulos Sepsis Neonatal Tempranaverock88No ratings yet
- Labor Induction Vs Expectant Management of Lowrisk Pregnancy 2018Document3 pagesLabor Induction Vs Expectant Management of Lowrisk Pregnancy 2018Maria MogosNo ratings yet
- Par EnteralDocument4 pagesPar EnteralshalukiriNo ratings yet
- tmp42AA TMPDocument8 pagestmp42AA TMPFrontiersNo ratings yet
- Research Article: Comparative Assessment of Cytokine Pattern in Early and Late Onset of Neonatal SepsisDocument9 pagesResearch Article: Comparative Assessment of Cytokine Pattern in Early and Late Onset of Neonatal SepsisPutri HolmesNo ratings yet
- Transfusions and Neurodevelopmental Outcomes in Extremely Low Gestation Neonates Enrolled in The PENUT Trial: A Randomized Clinical TrialDocument8 pagesTransfusions and Neurodevelopmental Outcomes in Extremely Low Gestation Neonates Enrolled in The PENUT Trial: A Randomized Clinical Trialdarcyelle666No ratings yet
- Review ArticleDocument8 pagesReview ArticleIdmNo ratings yet
- Early Childhood Diarrhea Predicts Cognitive Delays in Later Childhood Independently of MalnutritionDocument7 pagesEarly Childhood Diarrhea Predicts Cognitive Delays in Later Childhood Independently of MalnutritionYatil HidayaniNo ratings yet
- Colour of Meconium ND Il6Document4 pagesColour of Meconium ND Il6Akula KapilNo ratings yet
- Wattegedera 2008Document8 pagesWattegedera 2008Ignacio GualNo ratings yet
- Neonatal Group A Streptococcus Meningitis. Case Report and Literature ReviewDocument6 pagesNeonatal Group A Streptococcus Meningitis. Case Report and Literature ReviewJessyMomoNo ratings yet
- BUENISIMO Spittle2018 Prematuros HineDocument12 pagesBUENISIMO Spittle2018 Prematuros Hinejulianamd87No ratings yet
- ORATORIO Ocrelizumab EMPPDocument12 pagesORATORIO Ocrelizumab EMPPalmarazneurologiaNo ratings yet
- Full Text Jurnal LactobasilusDocument10 pagesFull Text Jurnal Lactobasilusrisda aulia putriNo ratings yet
- Minimising Neonatal Brain Injury: How Research in The Past Five Years Has Changed My Clinical PracticeDocument5 pagesMinimising Neonatal Brain Injury: How Research in The Past Five Years Has Changed My Clinical PracticeDian NovitasariNo ratings yet
- HHV-6 Specific T-Cell Immunity in Healthy Children and AdolescentsDocument8 pagesHHV-6 Specific T-Cell Immunity in Healthy Children and AdolescentsRahmad SyamsulNo ratings yet
- Early Enteral Feeding and Nosocomial Sepsis in Very Low Birthweight InfantsDocument4 pagesEarly Enteral Feeding and Nosocomial Sepsis in Very Low Birthweight InfantsaripbudiboyNo ratings yet
- Preimplantation Genetic Diagnosis For Aneuploidy Screening in Patients With Unexplained Recurrent MiscarriagesDocument5 pagesPreimplantation Genetic Diagnosis For Aneuploidy Screening in Patients With Unexplained Recurrent MiscarriagesAkmilia VeraNo ratings yet
- Preterm Labour, Antibiotics, and Cerebral Palsy: Scientific Impact Paper No. 33Document8 pagesPreterm Labour, Antibiotics, and Cerebral Palsy: Scientific Impact Paper No. 33oscarNo ratings yet
- Jurnal 8Document4 pagesJurnal 8Yoan YasrilNo ratings yet
- TMP FDE0Document5 pagesTMP FDE0FrontiersNo ratings yet
- Tmp305e TMPDocument3 pagesTmp305e TMPFrontiersNo ratings yet
- Main AbrDocument3 pagesMain AbrIndraWijayaNo ratings yet
- Clinical Conditions Associated With Post Natal Growth Failure in Preterm InfantsDocument1 pageClinical Conditions Associated With Post Natal Growth Failure in Preterm Infantsrubenporto5579No ratings yet
- Canine Fetus Immune System at Late DevelopmentDocument4 pagesCanine Fetus Immune System at Late DevelopmentWillNo ratings yet
- The Evolution of Primitive Reflexes in Extremely Premature InfantsDocument6 pagesThe Evolution of Primitive Reflexes in Extremely Premature InfantsPriydarshni KohliNo ratings yet
- Ref 5Document11 pagesRef 5Tiago BaraNo ratings yet
- Thy Box Timo Munoz-Chapuli2015Document6 pagesThy Box Timo Munoz-Chapuli2015PABLO ALBERTO SANCHEZ BARRERANo ratings yet
- Efficacy of Surfactant at Different Gestational Ages For Infants With Respiratory Distress Syndrome PDFDocument7 pagesEfficacy of Surfactant at Different Gestational Ages For Infants With Respiratory Distress Syndrome PDFnisaNo ratings yet
- Shachar2014 PDFDocument1 pageShachar2014 PDFMahlina Nur LailiNo ratings yet
- The Problems of Moderate Preterm Infants: Seminars in PerinatologyDocument4 pagesThe Problems of Moderate Preterm Infants: Seminars in Perinatologysam crushNo ratings yet
- Operative Management of Cryptorchidism Guidelines and RealityDocument9 pagesOperative Management of Cryptorchidism Guidelines and RealityRichard QiuNo ratings yet
- Laparoscopic Distal Splenoadrenal ShuntDocument5 pagesLaparoscopic Distal Splenoadrenal ShuntRichard QiuNo ratings yet
- Operative Management of Cryptorchidism Guidelines and RealityDocument9 pagesOperative Management of Cryptorchidism Guidelines and RealityRichard QiuNo ratings yet
- Chapter 151. Manual Testicular DetorsionDocument7 pagesChapter 151. Manual Testicular DetorsionRichard QiuNo ratings yet
- Nutrition-Digestion Vocabulary Study Guide - Unit 5Document3 pagesNutrition-Digestion Vocabulary Study Guide - Unit 5WordManInAthensNo ratings yet
- Relief For IBS, Cholitis, Crohn's and MoreDocument2 pagesRelief For IBS, Cholitis, Crohn's and MoreTheVitaminStore.comNo ratings yet
- Digestive SystemDocument2 pagesDigestive SystemReginaNo ratings yet
- Electroacupuctura VollDocument32 pagesElectroacupuctura Vollminunat50% (2)
- Chapter 11 Alimentary CanalDocument50 pagesChapter 11 Alimentary CanalLiana Du PlessisNo ratings yet
- Factors Affecting Drug AbsorptionDocument48 pagesFactors Affecting Drug Absorptionprashil charkariNo ratings yet
- Pig Anatomy PosterDocument1 pagePig Anatomy PosterBrandon Alexis Calderón AvellanedaNo ratings yet
- The Digestive SystemDocument2 pagesThe Digestive SystemAutumn GarofolaNo ratings yet
- Digestibility Determination in Fish Using Chromic Oxide Marking and Analysis of Contents From Different Segments of The Gastrointestinal TractDocument8 pagesDigestibility Determination in Fish Using Chromic Oxide Marking and Analysis of Contents From Different Segments of The Gastrointestinal TractRafael Hamakawa ViannaNo ratings yet
- Poultry BrochureDocument12 pagesPoultry BrochureWayanad AyurvedaNo ratings yet
- Li 020111 SPDocument12 pagesLi 020111 SPMonty J. RenovNo ratings yet
- Digestive System Background and ColoringDocument4 pagesDigestive System Background and ColoringVictoria LowmanNo ratings yet
- Frog Dissection WorksheetDocument7 pagesFrog Dissection WorksheetJoanna AcuñaNo ratings yet
- Detailed Lesson Plan in Science and Health 4 Unit 1 The Digestive SystemDocument6 pagesDetailed Lesson Plan in Science and Health 4 Unit 1 The Digestive Systemsheryl tianaNo ratings yet
- Parenteral and Enteral Nutrition in NeonateDocument22 pagesParenteral and Enteral Nutrition in NeonateAnonymous N2PHMnTIYLNo ratings yet
- Clinical Practice Guidelines For Allied Hirschsprung's Diseases - Practical Version PDFDocument89 pagesClinical Practice Guidelines For Allied Hirschsprung's Diseases - Practical Version PDFRudyard Paul AmistadNo ratings yet
- Embryology GIT 1-1Document48 pagesEmbryology GIT 1-1gtaha80No ratings yet
- Healing Codes: Grigori GrabovoiDocument47 pagesHealing Codes: Grigori GrabovoiPanther MelchizedekNo ratings yet
- Science Year 8 DietDocument5 pagesScience Year 8 DietJelle MustefaNo ratings yet
- Crohn's Disease Seminar FinalDocument27 pagesCrohn's Disease Seminar Finalshahad alshareefNo ratings yet
- Foot&Hand Map ReflexoDocument6 pagesFoot&Hand Map Reflexomusa_cristianNo ratings yet
- Kevin Yocum Digestive System 1Document39 pagesKevin Yocum Digestive System 1HomeyO'StasisNo ratings yet
- Brochure Pigeon (PDFDrive)Document104 pagesBrochure Pigeon (PDFDrive)Dummy DonNo ratings yet
- Clinical Chemistry of StoolDocument10 pagesClinical Chemistry of StoolbnmjgcNo ratings yet
- Cambridge O Level: BIOLOGY 5090/22Document20 pagesCambridge O Level: BIOLOGY 5090/22Aosaf Ershad ChowdhuryNo ratings yet
- Your Dynamic Profile: Wellness ReportDocument17 pagesYour Dynamic Profile: Wellness Reportvica sibueaNo ratings yet
- A Case Study On Typhoid FeverDocument42 pagesA Case Study On Typhoid FeverAna92% (13)
- Science4 - Q2 - Mod1 - Major Organs of The Human Body-ABRIDGEDDocument8 pagesScience4 - Q2 - Mod1 - Major Organs of The Human Body-ABRIDGEDRonie PadlanNo ratings yet
- Giardia LambliaDocument19 pagesGiardia LambliaAjishasughiNo ratings yet
- LEONCITO - Module 16 Worksheets 2Document12 pagesLEONCITO - Module 16 Worksheets 2Anton LeoncitoNo ratings yet