You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- 11-01-2022 El Rombo de La NFPADocument2 pages11-01-2022 El Rombo de La NFPALeydi LopezNo ratings yet
- Ultima Entrega Estadistica InferencialDocument4 pagesUltima Entrega Estadistica InferencialNicolas Pedraza100% (1)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Alimentación en Las Diferentes Etapas de La Vida ScribbDocument52 pagesAlimentación en Las Diferentes Etapas de La Vida Scribbsyoung2271% (7)
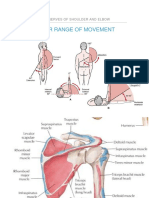
- Upperlimb Part 2Document59 pagesUpperlimb Part 2Jihuhn YuNo ratings yet
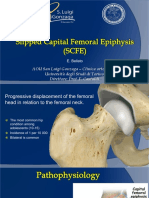
- 02-2. Pediatric-SCFEDocument23 pages02-2. Pediatric-SCFEJihuhn YuNo ratings yet
- 4 - Trunk Part 1Document32 pages4 - Trunk Part 1Jihuhn YuNo ratings yet
- Lower Limb - 18 - 10 - 2021Document39 pagesLower Limb - 18 - 10 - 2021Jihuhn YuNo ratings yet
- Ranking 2021 Per UniversitàDocument76 pagesRanking 2021 Per UniversitàJihuhn YuNo ratings yet
- Basic Cardiovascular Physiology-Pasquale Pagliaro, Raffaella RastaldoDocument438 pagesBasic Cardiovascular Physiology-Pasquale Pagliaro, Raffaella RastaldoJihuhn YuNo ratings yet
- Traguardo Cils b2 Pagine InizialiDocument6 pagesTraguardo Cils b2 Pagine InizialiJihuhn Yu100% (1)
- 02-1.WHO 2022-PitNETDocument21 pages02-1.WHO 2022-PitNETJihuhn YuNo ratings yet
- CILS B2 Modello SoluzioniDocument8 pagesCILS B2 Modello SoluzioniJihuhn YuNo ratings yet
- CZ120 (CW612N) : Technical DatasheetDocument1 pageCZ120 (CW612N) : Technical DatasheetAmrut KanungoNo ratings yet
- 1-SISTEMA DE DETECCIÓN DE ARCO ELÉCTRICO-V1-nuevoDocument5 pages1-SISTEMA DE DETECCIÓN DE ARCO ELÉCTRICO-V1-nuevoFrancisco DiazNo ratings yet
- Diagnostico Diferencial de Las ConvulsionesDocument5 pagesDiagnostico Diferencial de Las Convulsiones¡KyoNo ratings yet
- Actualizaciòn y Revisiòn de Las Normas de Los Servicios de Alimentación y Dietética HospitalariaDocument74 pagesActualizaciòn y Revisiòn de Las Normas de Los Servicios de Alimentación y Dietética HospitalariaXiomara BenavidesNo ratings yet
- Artículos de Opinión Versión Modelo de Rosa Montero 7Document39 pagesArtículos de Opinión Versión Modelo de Rosa Montero 7celialopez23mNo ratings yet
- Case Study 01: Food Shortage:: 2 Sem SY2018-19 Midterm Exam SUMMER/ Mdi/ Page - 1Document29 pagesCase Study 01: Food Shortage:: 2 Sem SY2018-19 Midterm Exam SUMMER/ Mdi/ Page - 1Marlyn IglesiasNo ratings yet
- Naparstek, F. Los Tres Tiempos de Freud (Teórico 4)Document12 pagesNaparstek, F. Los Tres Tiempos de Freud (Teórico 4)Lipu BolliniNo ratings yet
- Divisão de Grupos de Orientação de Estágio - NoturnoDocument13 pagesDivisão de Grupos de Orientação de Estágio - Noturnosillmara.sillvaoliveiraNo ratings yet
- Nist CSF 2020Document67 pagesNist CSF 2020jamssuperNo ratings yet
- Fruits 75 (6), 281-287 Effect of Fertilizer Treatments On Fruit Nutritional Quality of Plantain Cultivars and Derived HybridsDocument7 pagesFruits 75 (6), 281-287 Effect of Fertilizer Treatments On Fruit Nutritional Quality of Plantain Cultivars and Derived HybridsSol InvictusNo ratings yet
- Tipologia Gaston BergerDocument3 pagesTipologia Gaston BergerAlberto Rodriguez AlemanNo ratings yet
- Cuadernillo Pie Lenguaje 2021 IbDocument12 pagesCuadernillo Pie Lenguaje 2021 IbMaria EsterNo ratings yet
- Prudential Siem Reap Info Pack Content Final 25feb2020Document18 pagesPrudential Siem Reap Info Pack Content Final 25feb2020Venus ChuaNo ratings yet
- Trabalho-Ensaio Académico - DERL1Document17 pagesTrabalho-Ensaio Académico - DERL1Eveline PauloNo ratings yet
- Wran WraqDocument2 pagesWran WraqJanoNo ratings yet
- Toledo Power Company: Terms of ReferenceDocument8 pagesToledo Power Company: Terms of Referencejet toledo100% (1)
- Product Catalog: Ductless Mini-Splits, Light Commercial and Multi-Zone SystemsDocument72 pagesProduct Catalog: Ductless Mini-Splits, Light Commercial and Multi-Zone SystemsFernando ChaddadNo ratings yet
- Taller # 2 Del Curso Complementario de BioseguridadDocument3 pagesTaller # 2 Del Curso Complementario de BioseguridadDavid CabarcasNo ratings yet
- Cuadriláteros y Triángulos DorsoDocument1 pageCuadriláteros y Triángulos DorsoNadiaNo ratings yet
- Informe 1 Tecnología de LechesDocument5 pagesInforme 1 Tecnología de LechesDaisuke NuñezNo ratings yet
- Jurnal ProductGuide PDFDocument255 pagesJurnal ProductGuide PDFPrawira Ajie NNo ratings yet
- 15890Document15 pages15890uddinnadeemNo ratings yet
- Mitos, Leyendas y CuentosDocument16 pagesMitos, Leyendas y CuentosPaola BrazeiroNo ratings yet
- Grasas y AceitesDocument4 pagesGrasas y AceitesGINA SOFIA AMAGUA GONZALEZNo ratings yet
- Guia de Orientacion Modulo de Diseno de Sistemas de Manejo de Impacto Ambiental Saber Pro 2015 2 PDFDocument13 pagesGuia de Orientacion Modulo de Diseno de Sistemas de Manejo de Impacto Ambiental Saber Pro 2015 2 PDFsamiambiental2009No ratings yet
- Via Aerea ClasificacionesDocument21 pagesVia Aerea ClasificacionesQiqe CrNo ratings yet
- Practica NDocument18 pagesPractica Nvilmamarina100% (2)