You might also like
- Transfer Factors: Properties, Mechanism of Action and Its Clinical ApplicationsFrom EverandTransfer Factors: Properties, Mechanism of Action and Its Clinical ApplicationsNo ratings yet
- The Immunomodulatory Activities of Picria Fel-TerrDocument5 pagesThe Immunomodulatory Activities of Picria Fel-TerrMuhammad Fauzan LubisNo ratings yet
- Preclinical Immunology and Microbiology Review 2023: For USMLE Step 1 and COMLEX-USA Level 1From EverandPreclinical Immunology and Microbiology Review 2023: For USMLE Step 1 and COMLEX-USA Level 1Rating: 5 out of 5 stars5/5 (1)
- Microorganisms 08 01175Document14 pagesMicroorganisms 08 01175Karima El-yagoubiNo ratings yet
- Lipid MediatorsFrom EverandLipid MediatorsFiona M. CunninghamNo ratings yet
- Mco 12 06 0533Document8 pagesMco 12 06 0533Anonymous Wcj4C3jNo ratings yet
- Ahmed 2006Document9 pagesAhmed 2006Tharuka ChandrasiriNo ratings yet
- Ayissiowona2013 JQ1Document7 pagesAyissiowona2013 JQ1iforaNo ratings yet
- The Effect Of The Supernatant Of Coley'S Mixed Bacterial Toxin (Mbt) And Bacterial Lipopolysaccharide (Lps) On Serum Levels Of Tnf-Α, Il-12, And Vegf In MiceDocument5 pagesThe Effect Of The Supernatant Of Coley'S Mixed Bacterial Toxin (Mbt) And Bacterial Lipopolysaccharide (Lps) On Serum Levels Of Tnf-Α, Il-12, And Vegf In MiceIOSR Journal of PharmacyNo ratings yet
- Effects of Collagen IV On Neuroblastoma Cell Matrix-Related FunctionsDocument9 pagesEffects of Collagen IV On Neuroblastoma Cell Matrix-Related FunctionsdjackapNo ratings yet
- (Kyung Hee, Et Al, 2006) PGDocument4 pages(Kyung Hee, Et Al, 2006) PGWinda AlpiniawatiNo ratings yet
- Evaluation of Classical, Alternative, and Regulatory Functions of Bone Marrow-Derived MacrophagesDocument11 pagesEvaluation of Classical, Alternative, and Regulatory Functions of Bone Marrow-Derived MacrophagesgugicevdzoceNo ratings yet
- Moxibustion Activates Host Defense Against Herpes Simplex Virus Type I Through Augmentation of Cytokine ProductionDocument7 pagesMoxibustion Activates Host Defense Against Herpes Simplex Virus Type I Through Augmentation of Cytokine ProductionpedroNo ratings yet
- Liu 2005Document7 pagesLiu 2005Javier Cerda InfanteNo ratings yet
- Mitake Mushroom 3Document6 pagesMitake Mushroom 3dayra maría ixa marín salamancaNo ratings yet
- Hoat Tinh Cua AlbaconolDocument8 pagesHoat Tinh Cua Albaconolngoclinh87No ratings yet
- Flavonoids From Boldoa Purpurascens Inhibit ProinflammatoryDocument5 pagesFlavonoids From Boldoa Purpurascens Inhibit ProinflammatoryJOHN HELIBERTH ANDREY TORRES GRANADOSNo ratings yet
- Leucine 42Document8 pagesLeucine 42jdg487No ratings yet
- Kumolosasi E Et Al 2013 - Kinetics of Pro Inflamatory Cytokines in Response To LPSDocument8 pagesKumolosasi E Et Al 2013 - Kinetics of Pro Inflamatory Cytokines in Response To LPSeri.pinedarNo ratings yet
- Weisser Et Al 2013Document15 pagesWeisser Et Al 2013marcscianNo ratings yet
- BioFactors - 2010 - Bhui - Pineapple Bromelain Induces Autophagy Facilitating Apoptotic Response in Mammary CarcinomaDocument9 pagesBioFactors - 2010 - Bhui - Pineapple Bromelain Induces Autophagy Facilitating Apoptotic Response in Mammary CarcinomaFrancisco Salazar P.No ratings yet
- Plant Lectins Activate The NLRP3 Inflammasome To Promote Inflammatory DisordersDocument12 pagesPlant Lectins Activate The NLRP3 Inflammasome To Promote Inflammatory DisordersleonardopinheiroNo ratings yet
- Articulo IiDocument7 pagesArticulo IiOscarDavidGordilloGonzalezNo ratings yet
- Usnic-Acid Antiinflammatory PDFDocument5 pagesUsnic-Acid Antiinflammatory PDFRachel Lalaine Marie SialanaNo ratings yet
- NRP 9 E18Document6 pagesNRP 9 E18wawansiswadi01No ratings yet
- Ja 20066Document9 pagesJa 20066sivaNo ratings yet
- - Chen liang 2009 Signal-transducing mechanisms of ketamine-caused inhibition of interleukin-1β gene expression in lipopolysaccharide-stimulated murine macrophage-like Raw 264 PDFDocument11 pages- Chen liang 2009 Signal-transducing mechanisms of ketamine-caused inhibition of interleukin-1β gene expression in lipopolysaccharide-stimulated murine macrophage-like Raw 264 PDFRizalNo ratings yet
- Calprotectin Release From Human Neutrophils Is Induced by Porphyromonas Gingivalis Lipopolysaccharide Via The CD-14-Toll-like Receptor-Nuclear Factor JB PathwayDocument8 pagesCalprotectin Release From Human Neutrophils Is Induced by Porphyromonas Gingivalis Lipopolysaccharide Via The CD-14-Toll-like Receptor-Nuclear Factor JB PathwayCristianNo ratings yet
- Translational Studies - ABSTRACTS: G B A G B ADocument1 pageTranslational Studies - ABSTRACTS: G B A G B ASrujanKumarNo ratings yet
- 2 - Alterativas A Los Antibioticos (Ingles)Document21 pages2 - Alterativas A Los Antibioticos (Ingles)luis mozoNo ratings yet
- Tanshinone IIA Harmonizes The Crosstalk of Autophagy and 2019 InternationalDocument12 pagesTanshinone IIA Harmonizes The Crosstalk of Autophagy and 2019 InternationalfirdaNo ratings yet
- Journal of EthnopharmacologyDocument9 pagesJournal of EthnopharmacologyKaryn Olascuaga-CastilloNo ratings yet
- Protective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4Document7 pagesProtective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4harumiaikoNo ratings yet
- Role of Progesterone in Tlr4-Myd88-Dependent Signaling Pathway in Pre-EclampsiaDocument5 pagesRole of Progesterone in Tlr4-Myd88-Dependent Signaling Pathway in Pre-EclampsiaCarl Enrique PSNo ratings yet
- Protective Effect of 5-Hydroxy-3, 4, 7-Trimethoxyflavone Against Inflammation Induced by Lipopolysaccharide in RAW 264.7 Macrophage: in Vitro Study and in Silico ValidationDocument14 pagesProtective Effect of 5-Hydroxy-3, 4, 7-Trimethoxyflavone Against Inflammation Induced by Lipopolysaccharide in RAW 264.7 Macrophage: in Vitro Study and in Silico ValidationsopmaNo ratings yet
- Umar 2014Document11 pagesUmar 2014Andiaisyah macorawalieNo ratings yet
- Ketamine in ImunosupressanDocument6 pagesKetamine in ImunosupressanAnonymous MCduVEXNo ratings yet
- 6.hyun Kyung. en InglesDocument6 pages6.hyun Kyung. en InglesMAGALI GONZÁLEZ RODRÍGUEZNo ratings yet
- Cellular Immunology: Rupesh Kumar Shreewastav, Riyasat Ali, Jayaprakash Babu Uppada, D.N. RaoDocument8 pagesCellular Immunology: Rupesh Kumar Shreewastav, Riyasat Ali, Jayaprakash Babu Uppada, D.N. RaoDiego TulcanNo ratings yet
- Prolactin Increase Frecuency of T Helper FolicularDocument15 pagesProlactin Increase Frecuency of T Helper Folicularauroragamez2015No ratings yet
- 1 s2.0 S0955286316303722 MainDocument37 pages1 s2.0 S0955286316303722 MainA'in Qurrota A'yuninNo ratings yet
- 5570796Document9 pages5570796Leroy Christy LawalataNo ratings yet
- Popov 2002Document7 pagesPopov 2002DekdesNo ratings yet
- Phyt PDFDocument11 pagesPhyt PDFWACHIRACHAI PABUPRAPAPNo ratings yet
- Wang 1996Document7 pagesWang 1996Anh NguyenNo ratings yet
- 10 5798-Dicletip 1086124-2303017Document9 pages10 5798-Dicletip 1086124-2303017Rawaz Mahammad RasulNo ratings yet
- IDO1Document13 pagesIDO1Victor HaidarNo ratings yet
- Fan2016 Tissue Repair and Regeneration PDFDocument15 pagesFan2016 Tissue Repair and Regeneration PDFDavid LuNo ratings yet
- 1 s2.0 S0021925820870148 MainDocument11 pages1 s2.0 S0021925820870148 MainAina SarahNo ratings yet
- Macrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin ReceptorsDocument8 pagesMacrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin Receptorsvanessa_murillo_5No ratings yet
- Ijmm 25 4 593 PDFDocument8 pagesIjmm 25 4 593 PDFNickNo ratings yet
- Crozza 2020-BioRxiv-ENPP1 Inhibitors - ReviewDocument37 pagesCrozza 2020-BioRxiv-ENPP1 Inhibitors - ReviewBrahmam PujalaNo ratings yet
- Food Chemistry: Ana Cardeno, Marina Sanchez-Hidalgo, Marina Aparicio-Soto, Catalina Alarcón-de-la-LastraDocument7 pagesFood Chemistry: Ana Cardeno, Marina Sanchez-Hidalgo, Marina Aparicio-Soto, Catalina Alarcón-de-la-LastraSarah EmidioNo ratings yet
- Melendres Chem-103 Final-Requirements May-2022Document31 pagesMelendres Chem-103 Final-Requirements May-2022Bianca MelendresNo ratings yet
- Wang Et Al-2012-Journal of Pineal ResearchDocument8 pagesWang Et Al-2012-Journal of Pineal ResearchDionisio MesyeNo ratings yet
- Di 2017Document10 pagesDi 2017V LjNo ratings yet
- Ac MonoclonalDocument7 pagesAc MonoclonalRobert Larry Cordova DiazNo ratings yet
- Phosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersDocument11 pagesPhosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersMohammed Elias AlamNo ratings yet
- Effet Glucane Reishi 19 Janv 2023Document8 pagesEffet Glucane Reishi 19 Janv 2023ilyNo ratings yet
- Neutrophil Function in Small Animals: Shannon Jones HostetterDocument15 pagesNeutrophil Function in Small Animals: Shannon Jones HostetterNejraNo ratings yet
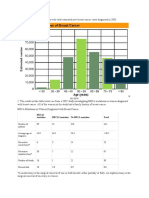
- Brca1 Mutation BRCA2 Mutation No BRCA Mutation Total: © 2011 FLVSDocument3 pagesBrca1 Mutation BRCA2 Mutation No BRCA Mutation Total: © 2011 FLVSJasmine VeraNo ratings yet
- Earth and Life Science Quarter 2 Module 4 FinalDocument14 pagesEarth and Life Science Quarter 2 Module 4 FinalNaima Kahlen100% (1)
- TCS Book RevisedDocument40 pagesTCS Book RevisedkkNo ratings yet
- IISC Bangalore PH.D Entrance Question Paper 2009Document22 pagesIISC Bangalore PH.D Entrance Question Paper 2009Abhay KumarNo ratings yet
- Bacara - JAOAC Vol 95 2012 - 508bDocument6 pagesBacara - JAOAC Vol 95 2012 - 508bDanaTNo ratings yet
- Identification of Malassezia SpeciesDocument3 pagesIdentification of Malassezia SpeciesSJ IraaNo ratings yet
- Ganoderma Lucidum A Traditional Chinese Medicine UDocument13 pagesGanoderma Lucidum A Traditional Chinese Medicine UTuan Anh LeNo ratings yet
- J. Nutr.-1997-Varga-819S-23SDocument5 pagesJ. Nutr.-1997-Varga-819S-23SVictor GilNo ratings yet
- Partial AutopsiesDocument5 pagesPartial AutopsiesTifano Prasali ArianNo ratings yet
- March Rqs COMPILED BY DR. STRANGEDocument89 pagesMarch Rqs COMPILED BY DR. STRANGEghassan100% (1)
- Psychopathology: Biological Basis of Behavior DisordersDocument9 pagesPsychopathology: Biological Basis of Behavior DisordersargatuNo ratings yet
- PCR Troubleshooting Guide 2Document2 pagesPCR Troubleshooting Guide 2Jagadeesh SelvamNo ratings yet
- Harding Et Al-2017-Movement DisordersDocument9 pagesHarding Et Al-2017-Movement DisordersNeryHdezNo ratings yet
- ANEMIA EtiologyDocument23 pagesANEMIA EtiologyAbhishek BhandariNo ratings yet
- Ilovepdf.comDocument348 pagesIlovepdf.comJeffrey AndersonNo ratings yet
- HEMOSTASISDocument6 pagesHEMOSTASIScatuiraneljhayyyNo ratings yet
- 16Document5 pages16dfngjlnNo ratings yet
- AbstractDocument4 pagesAbstractDinindu SiriwardeneNo ratings yet
- The Real Science Behind Why The COVID Vaccines Are Destroying Organ Function - Dangers of The Spike ProteinDocument40 pagesThe Real Science Behind Why The COVID Vaccines Are Destroying Organ Function - Dangers of The Spike ProteinMara Rebeka Herzog100% (7)
- 2014.MSG - Card Muscle Contr - Student QuestDocument2 pages2014.MSG - Card Muscle Contr - Student Questazuma12345No ratings yet
- Validation of The Brief Shame and Guilt Questionnaire For ChildrenDocument5 pagesValidation of The Brief Shame and Guilt Questionnaire For ChildrenVornicu TatianNo ratings yet
- Insider's Guide 37Document18 pagesInsider's Guide 37whiteyasminNo ratings yet
- Veterinary ParasitologyDocument4 pagesVeterinary Parasitologysimran kaurNo ratings yet
- Myopia:: A Modern Yet Reversible DiseaseDocument22 pagesMyopia:: A Modern Yet Reversible Diseasedr_vuja100% (1)
- 4612 4621 Vitamin D Supplementation For Osteoporosis in Older AdultsDocument10 pages4612 4621 Vitamin D Supplementation For Osteoporosis in Older AdultsMohammedNo ratings yet
- TY BSC Microbiology PDFDocument26 pagesTY BSC Microbiology PDFShubhamNo ratings yet
- Pathology Lecture 3, Cell Injury (Continued) (Lecture Notes)Document11 pagesPathology Lecture 3, Cell Injury (Continued) (Lecture Notes)Ali Al-Qudsi100% (3)
- Molecular Docking: in Computer Aided Drug DesignDocument26 pagesMolecular Docking: in Computer Aided Drug DesignGravelandNo ratings yet
- Freebirth LeseprobeDocument37 pagesFreebirth LeseprobedidibutterflyNo ratings yet
- Stress MGT - Chandana J. HewawasamDocument48 pagesStress MGT - Chandana J. HewawasamChandana J.HewawasamNo ratings yet
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeFrom EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeRating: 4.5 out of 5 stars4.5/5 (3)
- Meltdown: Nuclear disaster and the human cost of going criticalFrom EverandMeltdown: Nuclear disaster and the human cost of going criticalRating: 5 out of 5 stars5/5 (5)
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableFrom EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableRating: 3.5 out of 5 stars3.5/5 (22)
- Chemistry: a QuickStudy Laminated Reference GuideFrom EverandChemistry: a QuickStudy Laminated Reference GuideRating: 5 out of 5 stars5/5 (1)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolFrom EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolNo ratings yet
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsFrom EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsRating: 4 out of 5 stars4/5 (146)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeFrom EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeRating: 4 out of 5 stars4/5 (1)
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilFrom EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilRating: 5 out of 5 stars5/5 (1)
- The Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookFrom EverandThe Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookNo ratings yet
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsFrom EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsRating: 5 out of 5 stars5/5 (3)
- Handbook of Formulating Dermal Applications: A Definitive Practical GuideFrom EverandHandbook of Formulating Dermal Applications: A Definitive Practical GuideNo ratings yet
- Formulating, Packaging, and Marketing of Natural Cosmetic ProductsFrom EverandFormulating, Packaging, and Marketing of Natural Cosmetic ProductsNo ratings yet
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeFrom EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeRating: 5 out of 5 stars5/5 (1)
- Bioplastics: A Home Inventors HandbookFrom EverandBioplastics: A Home Inventors HandbookRating: 4 out of 5 stars4/5 (2)
- Phase Equilibria in Chemical EngineeringFrom EverandPhase Equilibria in Chemical EngineeringRating: 4 out of 5 stars4/5 (11)
- AP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeFrom EverandAP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeRating: 5 out of 5 stars5/5 (1)
- A Perfect Red: Empire, Espionage, and the Quest for the Color of DesireFrom EverandA Perfect Red: Empire, Espionage, and the Quest for the Color of DesireRating: 4 out of 5 stars4/5 (129)
- Transformer: The Deep Chemistry of Life and DeathFrom EverandTransformer: The Deep Chemistry of Life and DeathRating: 4.5 out of 5 stars4.5/5 (13)