You might also like
- Keys To The Ultimate Freedom PDFDocument245 pagesKeys To The Ultimate Freedom PDFdebtru100% (2)
- Focus 2011 WiringDocument139 pagesFocus 2011 Wiringphalksturm100% (1)
- Giordano Bruno and The RosicruciansDocument11 pagesGiordano Bruno and The RosicruciansGuido del GiudiceNo ratings yet
- ED 104 Module 1 Vision, Policy, Goal and Objectives of Special EducationDocument12 pagesED 104 Module 1 Vision, Policy, Goal and Objectives of Special EducationAngela Diaz100% (9)
- Idoc - Pub Medical Check Up FormDocument4 pagesIdoc - Pub Medical Check Up FormBobly SweaggerNo ratings yet
- 1 s2.0 S0014299921000522 MainDocument9 pages1 s2.0 S0014299921000522 MainMuhammad Reza FirdausNo ratings yet
- Outcomes in Patients With COVID-19 Infection Taking ACEI/ARBDocument4 pagesOutcomes in Patients With COVID-19 Infection Taking ACEI/ARBfk unswagatiNo ratings yet
- Risk of Sars-Cov-2 Infection and Covid-19 Prognosis With The Use of Renin - Angiotensin - Aldosterone System (Raas) Inhibitors: A Systematic ReviewDocument10 pagesRisk of Sars-Cov-2 Infection and Covid-19 Prognosis With The Use of Renin - Angiotensin - Aldosterone System (Raas) Inhibitors: A Systematic ReviewnembutalNo ratings yet
- Role of Angiotensin-Converting Enzyme 2 (ACE2) in COVID-19: Review Open AccessDocument10 pagesRole of Angiotensin-Converting Enzyme 2 (ACE2) in COVID-19: Review Open AccessjakpowerNo ratings yet
- EliabethDocument6 pagesEliabethmadelineNo ratings yet
- Cardiovascular System and COVID-19 - Manifestations and TherapeuticsDocument11 pagesCardiovascular System and COVID-19 - Manifestations and TherapeuticsmimiNo ratings yet
- The COVID-19 Pandemic: Critical Reviews in Clinical Laboratory SciencesDocument25 pagesThe COVID-19 Pandemic: Critical Reviews in Clinical Laboratory SciencesSir Ahmet AydınNo ratings yet
- 202 FullDocument6 pages202 FullMariana CabralNo ratings yet
- Ischemic Stroke in COVID-19-Positive Patients - An Overview of SARS-CoV-2 and Thrombotic Mechanisms For The NeurointerventionalistDocument6 pagesIschemic Stroke in COVID-19-Positive Patients - An Overview of SARS-CoV-2 and Thrombotic Mechanisms For The Neurointerventionalistarif 2006No ratings yet
- The COVID-19 PandemicDocument25 pagesThe COVID-19 PandemicJasper VanhaerenNo ratings yet
- 2021 Article 2395Document15 pages2021 Article 2395danyescalerapumas24No ratings yet
- A Hypothesis For Pathobiology and Treatment of COVID-19Document20 pagesA Hypothesis For Pathobiology and Treatment of COVID-19youssef arariNo ratings yet
- 1 s2.0 S0188440920302927 MainDocument5 pages1 s2.0 S0188440920302927 MainRheum RaponticumNo ratings yet
- Narrative Reviews: Diarrhea During COVID-19 Infection: Pathogenesis, Epidemiology, Prevention, and ManagementDocument10 pagesNarrative Reviews: Diarrhea During COVID-19 Infection: Pathogenesis, Epidemiology, Prevention, and ManagementWaqasKhanNo ratings yet
- Renin-Angiotensin System Blockers and The COVID-19 Pandemic: ReviewDocument4 pagesRenin-Angiotensin System Blockers and The COVID-19 Pandemic: ReviewClaudio Castro TrujilloNo ratings yet
- 2021 12 BijpsreviewDocument9 pages2021 12 BijpsreviewNguyên NguyễnNo ratings yet
- Epid Pubmed 1 (Bab2) .En - IdDocument11 pagesEpid Pubmed 1 (Bab2) .En - IdSHAFIYA ADELINANo ratings yet
- Excellent Paper On Covid 19Document20 pagesExcellent Paper On Covid 19GalenNo ratings yet
- Potential Harmful Effects of Discontinuing Ace-Inhibitors and Arbs in Covid-19 PatientsDocument8 pagesPotential Harmful Effects of Discontinuing Ace-Inhibitors and Arbs in Covid-19 PatientsmadelineNo ratings yet
- Estado Del Arte CoronavirusDocument17 pagesEstado Del Arte CoronavirusJosé AñorgaNo ratings yet
- Naguib - Potential Relationships Between COVID-19 and The Thyroid Gland An UpdateDocument16 pagesNaguib - Potential Relationships Between COVID-19 and The Thyroid Gland An UpdateerixNo ratings yet
- 13 ACE2 Receptor For Covid19Document11 pages13 ACE2 Receptor For Covid19mmaNo ratings yet
- Multisystem Imaging Manifesta-Tions of COVID-19, Part 2: From Cardiac Complications To Pediatric ManifestationsDocument27 pagesMultisystem Imaging Manifesta-Tions of COVID-19, Part 2: From Cardiac Complications To Pediatric ManifestationsAdiel OjedaNo ratings yet
- Diarrhea During COVID-19 InfectionDocument11 pagesDiarrhea During COVID-19 InfectionMan'SzAr'diAnSyAhNo ratings yet
- Comment: Covid-19 and The Cardiovascular SystemDocument2 pagesComment: Covid-19 and The Cardiovascular SystemplasmaNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateZâna ZorilorNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateRocio GmarNo ratings yet
- Drug Sars-CovDocument20 pagesDrug Sars-CovSilvi Zakiyatul IlmiyahNo ratings yet
- A Review On Sars Cov 2 Virology, Pathophysiology, Animal Models, and Anti Viral InterventionsDocument22 pagesA Review On Sars Cov 2 Virology, Pathophysiology, Animal Models, and Anti Viral InterventionsalodiarkNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateainindyaNo ratings yet
- The Cardiovascular Aspect of COVID 19Document11 pagesThe Cardiovascular Aspect of COVID 19Roberto Pulido SepulvedaNo ratings yet
- Consequences of COVID-19 For The PancreasDocument14 pagesConsequences of COVID-19 For The PancreasNormanNo ratings yet
- Breve Reseña Sobre Medicamentos y Vacunas Reutilizados para El Posible Tratamiento de COVID-19Document15 pagesBreve Reseña Sobre Medicamentos y Vacunas Reutilizados para El Posible Tratamiento de COVID-19Cristal Jewels DiamondNo ratings yet
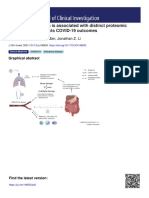
- SARS-CoV-2 Viremia Is Associated With Distinct Proteomic Pathways and Predicts COVID-19 OutcomesDocument13 pagesSARS-CoV-2 Viremia Is Associated With Distinct Proteomic Pathways and Predicts COVID-19 OutcomesYoga Nugraha WNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateArifin SiregarNo ratings yet
- Angiotensin InhibitionDocument3 pagesAngiotensin Inhibitionvivitri.dewiNo ratings yet
- Journal Pre-Proof: European Journal of Internal MedicineDocument30 pagesJournal Pre-Proof: European Journal of Internal MedicineNabil Sangga BuanaNo ratings yet
- Artigo 5Document5 pagesArtigo 5Paloma PaixãoNo ratings yet
- Articulo 3 - Disfuncion NeurologicaDocument7 pagesArticulo 3 - Disfuncion NeurologicaClaudia SalasNo ratings yet
- Sars-Cov-2 Receptor and Renin-Angiotensin System Regulation: Impact of Genetics Variants in Ace2 GeneDocument9 pagesSars-Cov-2 Receptor and Renin-Angiotensin System Regulation: Impact of Genetics Variants in Ace2 GeneInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateMaransdyka PurnamasidiNo ratings yet
- 1 s2.0 S2405844022021466 MainDocument8 pages1 s2.0 S2405844022021466 MainhogirimmunologyNo ratings yet
- 5400-Article Text-31589-2-10-20210820Document12 pages5400-Article Text-31589-2-10-20210820Voltaire Méndez RodríguezNo ratings yet
- Association Between ABO Blood Groups and Susceptibility To COVID 19Document4 pagesAssociation Between ABO Blood Groups and Susceptibility To COVID 19Arpana HazarikaNo ratings yet
- Laboratory Testing of Sars-Cov, Mers-Cov, and Sars-Cov-2 (2019-Ncov) : Current Status, Challenges, and CountermeasuresDocument14 pagesLaboratory Testing of Sars-Cov, Mers-Cov, and Sars-Cov-2 (2019-Ncov) : Current Status, Challenges, and CountermeasuresDanteraiantNo ratings yet
- Bosso Et Al 2020 Role of ACE2 Polymorphisms in HT and COVID19Document7 pagesBosso Et Al 2020 Role of ACE2 Polymorphisms in HT and COVID19Golden Era BookwormNo ratings yet
- Ssentogo Et Al.,2020Document7 pagesSsentogo Et Al.,2020madelineNo ratings yet
- 5 Emerging Diseases Corona Eng 2021Document46 pages5 Emerging Diseases Corona Eng 2021Vlad SmithNo ratings yet
- Immune Response Covid19Document9 pagesImmune Response Covid19Ario DaniantoNo ratings yet
- Knowing When and How To Fight: COVID-19 Between Viral Clearance and Immune ToleranceDocument11 pagesKnowing When and How To Fight: COVID-19 Between Viral Clearance and Immune ToleranceAri Trianto WibowoNo ratings yet
- Antihipertensive CovidDocument2 pagesAntihipertensive CovidIsa CretuNo ratings yet
- BBA - Molecular Basis of Disease: ReviewDocument12 pagesBBA - Molecular Basis of Disease: ReviewautomationenggNo ratings yet
- COVID 19 in The Heart and The Lungs: Could We "Notch" The Inflammatory Storm?Document8 pagesCOVID 19 in The Heart and The Lungs: Could We "Notch" The Inflammatory Storm?laax_siuyenNo ratings yet
- COVID-19: Pathophysiology, Diagnosis, Complications and Investigational TherapeuticsDocument8 pagesCOVID-19: Pathophysiology, Diagnosis, Complications and Investigational TherapeuticsHeidiNo ratings yet
- COVID para Cardiologistas JACC 2020Document19 pagesCOVID para Cardiologistas JACC 2020Maria Do SocorroNo ratings yet
- Therapi and Diagnostic CovidDocument22 pagesTherapi and Diagnostic CovidAni Rahayu0% (1)
- Pathogens: Biology and Pathogenesis of Sars-Cov-2: Understandings For Therapeutic Developments Against Covid-19Document27 pagesPathogens: Biology and Pathogenesis of Sars-Cov-2: Understandings For Therapeutic Developments Against Covid-19neftaliNo ratings yet
- Acei/Arb Therapy in Covid-19: The Double-Edged Sword of Ace2 and Sars-Cov-2 Viral DockingDocument3 pagesAcei/Arb Therapy in Covid-19: The Double-Edged Sword of Ace2 and Sars-Cov-2 Viral Dockingpond_1993No ratings yet
- An Update on SARS-CoV-2: Damage-response Framework, Potential Therapeutic Avenues and the Impact of Nanotechnology on COVID-19 TherapyFrom EverandAn Update on SARS-CoV-2: Damage-response Framework, Potential Therapeutic Avenues and the Impact of Nanotechnology on COVID-19 TherapyNo ratings yet
- Roztocil 2003Document9 pagesRoztocil 2003Anett Pappné LeppNo ratings yet
- International Angiology61 - 1989Document8 pagesInternational Angiology61 - 1989Anett Pappné LeppNo ratings yet
- Laurent 1988Document8 pagesLaurent 1988Anett Pappné LeppNo ratings yet
- Martinez Zapata - Et - Al 2016 .Sup 2Document172 pagesMartinez Zapata - Et - Al 2016 .Sup 2Anett Pappné LeppNo ratings yet
- Pokrovsky 2007Document12 pagesPokrovsky 2007Anett Pappné LeppNo ratings yet
- Pushpakom 2018Document18 pagesPushpakom 2018Anett Pappné LeppNo ratings yet
- Bao 2021Document11 pagesBao 2021Anett Pappné LeppNo ratings yet
- Jqac 039Document13 pagesJqac 039Anett Pappné LeppNo ratings yet
- Carlsonjones 2012Document7 pagesCarlsonjones 2012Anett Pappné LeppNo ratings yet
- Ijo 13 09 1443Document8 pagesIjo 13 09 1443Anett Pappné LeppNo ratings yet
- Brainsci 11 01111Document13 pagesBrainsci 11 01111Anett Pappné LeppNo ratings yet
- 2020 Article 642Document9 pages2020 Article 642Anett Pappné LeppNo ratings yet
- Innovative Injection Rate Control With Next Generation Common Rail Fuel Injection SystemDocument8 pagesInnovative Injection Rate Control With Next Generation Common Rail Fuel Injection SystemRakesh BiswasNo ratings yet
- Deep Learning Unit-1 FinalsDocument23 pagesDeep Learning Unit-1 FinalsAnonymous xMYE0TiNBcNo ratings yet
- User Manual HP Officejet 8210Document95 pagesUser Manual HP Officejet 8210ponidiNo ratings yet
- mathematicsproficiencyattitudeandperformanceofgrade9studentsinprivatehighschoolsinBukidnonPhilippines PDFDocument15 pagesmathematicsproficiencyattitudeandperformanceofgrade9studentsinprivatehighschoolsinBukidnonPhilippines PDFNacivah BasherNo ratings yet
- Media Literacy Dossier Duran Et Al 2008 SummarybyWangDocument2 pagesMedia Literacy Dossier Duran Et Al 2008 SummarybyWanglsemediapolicyNo ratings yet
- Industry CPD: Modelling Buildings For Seismic AnalysisDocument4 pagesIndustry CPD: Modelling Buildings For Seismic Analysisshare4learnNo ratings yet
- Regional Integration in Industrial Development in ECOWAS by Bitrus Kyangmah Samaila, BSC, MSC, MBA in View, University of Maiduguri, Nigeria.Document9 pagesRegional Integration in Industrial Development in ECOWAS by Bitrus Kyangmah Samaila, BSC, MSC, MBA in View, University of Maiduguri, Nigeria.BitrusNo ratings yet
- Learning From Others and Reviewing Literature: Zoila D. Espiritu, L.P.T., M.A.Ed. Stephanie P. MonteroDocument10 pagesLearning From Others and Reviewing Literature: Zoila D. Espiritu, L.P.T., M.A.Ed. Stephanie P. Monterochararat marnieNo ratings yet
- Oil Shows Description: (I) Sample Under Microscope (Under White Light)Document1 pageOil Shows Description: (I) Sample Under Microscope (Under White Light)deyaa SadoonNo ratings yet
- Consider The Rape of The Lock As A Social SatireDocument3 pagesConsider The Rape of The Lock As A Social SatireElena AiylaNo ratings yet
- River System of BangladeshDocument3 pagesRiver System of Bangladeshtonmoy50% (2)
- Knowage Baby Steps PDFDocument230 pagesKnowage Baby Steps PDFworkflow m3cNo ratings yet
- Catalogo PDFGeneratorDocument3 pagesCatalogo PDFGeneratoredwraulNo ratings yet
- IPC Lab 9Document12 pagesIPC Lab 9abdullah1996No ratings yet
- Harsh PatelDocument14 pagesHarsh PatelPradeepNo ratings yet
- Nemo Analyze: Professional Post-Processing of Drive Test DataDocument21 pagesNemo Analyze: Professional Post-Processing of Drive Test DataMohammed ShakilNo ratings yet
- 102 RadionicsDocument3 pages102 RadionicsResursaFun RoNo ratings yet
- How To Represent Geospatial Data in SDMX 20181022Document47 pagesHow To Represent Geospatial Data in SDMX 20181022scorpio1878No ratings yet
- s1 English Scope and Sequence ChecklistDocument18 pagess1 English Scope and Sequence Checklistapi-280236110No ratings yet
- Format For The Industrial Training ReportDocument7 pagesFormat For The Industrial Training ReportAbdulHalimNo ratings yet
- Maryada Purushotham RamDocument1 pageMaryada Purushotham RamNithin SridharNo ratings yet
- Ed 352649Document269 pagesEd 352649Hồng HạnhNo ratings yet
- Equipment ListDocument6 pagesEquipment ListJustin GordonNo ratings yet
- Ferrite Core Transformer Design Engineering and Technology 6Document10 pagesFerrite Core Transformer Design Engineering and Technology 6nedian_2006100% (1)
- Aceleradores Serie 6900Document2 pagesAceleradores Serie 6900jdavis5548No ratings yet