You might also like
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Complete Enochian HandbookDocument29 pagesThe Complete Enochian Handbook9toesroze100% (7)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- South Pointe Park LayoutDocument117 pagesSouth Pointe Park Layoutmary34d100% (1)
- Actividades Coordenadas GeograficasDocument4 pagesActividades Coordenadas GeograficasGraciela Alejandra Gómez Gutiérrez50% (2)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5810)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Biotechnology and Plant Breeding: Applications and Approaches for Developing Improved CultivarsFrom EverandBiotechnology and Plant Breeding: Applications and Approaches for Developing Improved CultivarsAluízio BorémRating: 5 out of 5 stars5/5 (3)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Der Yidam Und Seine Yogini - Und Dann Sprach ShaktiDocument68 pagesDer Yidam Und Seine Yogini - Und Dann Sprach ShaktiGauri B.No ratings yet
- Bouveresse Prodiges Et Vertiges de L AnalogieDocument79 pagesBouveresse Prodiges Et Vertiges de L Analogieiulian_1100% (1)
- Abiotic and Biotic Stresses in Soybean Production: Soybean Production Volume 1From EverandAbiotic and Biotic Stresses in Soybean Production: Soybean Production Volume 1Rating: 4 out of 5 stars4/5 (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Advances in Agricultural MicrobiologyFrom EverandAdvances in Agricultural MicrobiologyN.S. Subba RaoRating: 5 out of 5 stars5/5 (2)
- Greenhouse Gas Emissions From Aquaculture: A Life Cycle Assessment of Three Asian SystemsFrom EverandGreenhouse Gas Emissions From Aquaculture: A Life Cycle Assessment of Three Asian SystemsNo ratings yet
- Functional Diversity of Mycorrhiza and Sustainable Agriculture: Management to Overcome Biotic and Abiotic StressesFrom EverandFunctional Diversity of Mycorrhiza and Sustainable Agriculture: Management to Overcome Biotic and Abiotic StressesNo ratings yet
- Relatório de Estágio Obrigatório ElinaDocument24 pagesRelatório de Estágio Obrigatório ElinaCesar Botelho100% (1)
- Microbial Services in Restoration EcologyFrom EverandMicrobial Services in Restoration EcologyJay Shankar SinghNo ratings yet
- Expert Consultation on Nutrition Indicators for Biodiversity Food CompositionFrom EverandExpert Consultation on Nutrition Indicators for Biodiversity Food CompositionNo ratings yet
- Emerging Trends in Agri-Nanotechnology: Fundamental and Applied AspectsFrom EverandEmerging Trends in Agri-Nanotechnology: Fundamental and Applied AspectsNo ratings yet
- Biodiversity and Livelihood: Lessons from Community Research in IndiaFrom EverandBiodiversity and Livelihood: Lessons from Community Research in IndiaNo ratings yet
- A Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyFrom EverandA Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyNo ratings yet
- Molecular and Physiological Insights into Plant Stress Tolerance and Applications in Agriculture (Part 2)From EverandMolecular and Physiological Insights into Plant Stress Tolerance and Applications in Agriculture (Part 2)No ratings yet
- Biostimulants for Crop Production and Sustainable AgricultureFrom EverandBiostimulants for Crop Production and Sustainable AgricultureMirza HasanuzzamanNo ratings yet
- Microbes for Climate Resilient AgricultureFrom EverandMicrobes for Climate Resilient AgriculturePrem Lal KashyapNo ratings yet
- Voluntary Guidelines for the Conservation and Sustainable Use of Farmers' Varieties/LandracesFrom EverandVoluntary Guidelines for the Conservation and Sustainable Use of Farmers' Varieties/LandracesNo ratings yet
- Trends of Applied Microbiology for Sustainable EconomyFrom EverandTrends of Applied Microbiology for Sustainable EconomyRavindra SoniNo ratings yet
- Pacific Regional Synthesis for the State of the World’s Biodiversity for Food and AgricultureFrom EverandPacific Regional Synthesis for the State of the World’s Biodiversity for Food and AgricultureNo ratings yet
- Wheat and Barley Grain BiofortificationFrom EverandWheat and Barley Grain BiofortificationOm Prakash GuptaNo ratings yet
- The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture: FAO Commission on Genetic Resources for Food and Agriculture AssessmentsFrom EverandThe Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture: FAO Commission on Genetic Resources for Food and Agriculture AssessmentsNo ratings yet
- Risk Profile - Group B Streptococcus (GBS) – Streptococcus Agalactiae Sequence Type (ST) 283 in Freshwater FishFrom EverandRisk Profile - Group B Streptococcus (GBS) – Streptococcus Agalactiae Sequence Type (ST) 283 in Freshwater FishNo ratings yet
- Control and Manipulation of Animal Growth: Proceedings of Previous Easter Schools in Agricultural ScienceFrom EverandControl and Manipulation of Animal Growth: Proceedings of Previous Easter Schools in Agricultural ScienceNo ratings yet
- Voluntary Guidelines for the Conservation and Sustainable Use of Crop Wild Relatives and Wild Food PlantsFrom EverandVoluntary Guidelines for the Conservation and Sustainable Use of Crop Wild Relatives and Wild Food PlantsNo ratings yet
- Chemical Manipulation of Crop Growth and Development: Proceedings of Previous Easter Schools in Agricultural ScienceFrom EverandChemical Manipulation of Crop Growth and Development: Proceedings of Previous Easter Schools in Agricultural ScienceNo ratings yet
- Plant-Bacteria Interactions: Strategies and Techniques to Promote Plant GrowthFrom EverandPlant-Bacteria Interactions: Strategies and Techniques to Promote Plant GrowthNo ratings yet
- Imperatives for Improvement of Food Safety in Fruit and Vegetable Value Chains in Viet NamFrom EverandImperatives for Improvement of Food Safety in Fruit and Vegetable Value Chains in Viet NamNo ratings yet
- Tarea S 8 2023Document3 pagesTarea S 8 2023Ramón Manuel Minaya RodríguezNo ratings yet
- FO-QU-06 Solicitud de Acciones Correctivas y Mejora 2021Document2 pagesFO-QU-06 Solicitud de Acciones Correctivas y Mejora 2021Alix ValenciaNo ratings yet
- Mackelbert Sanchez 63751600 85331600 2019Document110 pagesMackelbert Sanchez 63751600 85331600 2019wijdane chniouliNo ratings yet
- Propagacion de IncertidumbresDocument26 pagesPropagacion de Incertidumbreseduardo100% (1)
- Informe de PM10Document20 pagesInforme de PM10Ebe LynNo ratings yet
- Segundo MomentoDocument9 pagesSegundo MomentoFran EscobarNo ratings yet
- Compreensão e Interpretação de TextoDocument4 pagesCompreensão e Interpretação de TextoandreaNo ratings yet
- Evaluacion de Filosofia Escenario 5 IdelverDocument6 pagesEvaluacion de Filosofia Escenario 5 IdelverAngie Liceth Lopez RamirezNo ratings yet
- Practicas ConcentracionDocument17 pagesPracticas ConcentracionMiguel RodriguezNo ratings yet
- U30 New TechnologiesDocument1 pageU30 New TechnologiesPaula SuciuNo ratings yet
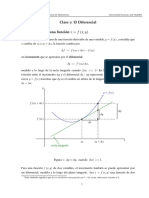
- Clase 07. El DiferencialDocument5 pagesClase 07. El DiferencialJhonatan Felipe Tovar RomeroNo ratings yet
- Beam Design For Moment, Shear & TorsionDocument6 pagesBeam Design For Moment, Shear & TorsionRajendra BadjatyaNo ratings yet
- 06.03.20. Central Hidroeléctrica Simón Bolívar. (GHC)Document3 pages06.03.20. Central Hidroeléctrica Simón Bolívar. (GHC)DarianaNo ratings yet
- Implementación de Buenas Prácticas de Promoción de Personal y Su Relación Con La Cultura y El Compromiso Con La OrganizaciónDocument2 pagesImplementación de Buenas Prácticas de Promoción de Personal y Su Relación Con La Cultura y El Compromiso Con La Organizacióncristelad200290No ratings yet
- Ciclo de La Tarea2 Anny MartinezDocument8 pagesCiclo de La Tarea2 Anny MartinezAnny Liseth Martinez SeguraNo ratings yet
- Lsbu Dissertation SubmissionDocument7 pagesLsbu Dissertation SubmissionWriteMyCollegePaperForMeSingapore100% (1)
- Proposal MELDocument14 pagesProposal MELkopicoloNo ratings yet
- Embarazo Adolescente. Paula FaisondDocument10 pagesEmbarazo Adolescente. Paula FaisondHunt KarenNo ratings yet
- Research Papers On Climate Change Global WarmingDocument4 pagesResearch Papers On Climate Change Global Warminggw1nm9nbNo ratings yet
- Informe U Oficio de CanalizaciónDocument3 pagesInforme U Oficio de CanalizaciónxochilNo ratings yet
- Informe N°9 Determinación de ClorurosDocument16 pagesInforme N°9 Determinación de ClorurosRonyNo ratings yet
- Preguntas de Razonamiento 1Document2 pagesPreguntas de Razonamiento 1Lola TellezNo ratings yet
- 35MW Solar Power Project - Upper West Regional Sites - Non Technical Summary ReportDocument109 pages35MW Solar Power Project - Upper West Regional Sites - Non Technical Summary ReportElijahNo ratings yet
- 599 2344 2 PBDocument6 pages599 2344 2 PBJhei QuintanaNo ratings yet