You might also like
- Perivascular Adipose Tissue 2023 JDocument8 pagesPerivascular Adipose Tissue 2023 Jmiguel contrerasNo ratings yet
- Agabiti Rosei 2023 Influence of Perivascular Adipose TDocument9 pagesAgabiti Rosei 2023 Influence of Perivascular Adipose Tmiguel contrerasNo ratings yet
- 17 PVAT (Gao)Document11 pages17 PVAT (Gao)Júlia Martins F. de SouzaNo ratings yet
- Vascular Endothelium e Gatekeeper of Vessel Health: AtherosclerosisDocument17 pagesVascular Endothelium e Gatekeeper of Vessel Health: AtherosclerosisLia FitriaNo ratings yet
- 10 1016@j Redox 2017 01 007Document17 pages10 1016@j Redox 2017 01 007yalocim666No ratings yet
- Disfunctional Vascular Endothelium As A Driver of Atherosclerosis - Emergind Insights Into Pathogenesis and Treatment 2021Document18 pagesDisfunctional Vascular Endothelium As A Driver of Atherosclerosis - Emergind Insights Into Pathogenesis and Treatment 2021Camila VidigalNo ratings yet
- FP Ago 2020 - Mecanismos Moleculares y Mecánicos Que Regulan El Cierre Del Ductus en PrematurosDocument19 pagesFP Ago 2020 - Mecanismos Moleculares y Mecánicos Que Regulan El Cierre Del Ductus en Prematurosjulius castelloNo ratings yet
- 1 s2.0 S2666667722000058 MainDocument14 pages1 s2.0 S2666667722000058 Mainmiskica miskicaNo ratings yet
- Angiogenesis - Molecular Mechanisms and Clinical Applications of AngiogenesisDocument28 pagesAngiogenesis - Molecular Mechanisms and Clinical Applications of AngiogenesisCher IshNo ratings yet
- It Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and DiseaseDocument14 pagesIt Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and Diseasesri karuniaNo ratings yet
- Nutrients 13 02603 v2Document18 pagesNutrients 13 02603 v2David CampoverdeNo ratings yet
- Vasculita Shavit2018Document9 pagesVasculita Shavit2018JasNo ratings yet
- 04 Relatic CE PDFDocument13 pages04 Relatic CE PDFSang Ayu Suryantari PpiNo ratings yet
- Nihms 1873457Document15 pagesNihms 1873457Hanssel Viramontes CastroNo ratings yet
- TMP AED3Document13 pagesTMP AED3FrontiersNo ratings yet
- Fluid Therapy and The Microcirculation in Health and Critical IllnessDocument11 pagesFluid Therapy and The Microcirculation in Health and Critical IllnessAdote DrmNo ratings yet
- Engineering Grafts: Genetic Vein Resistant AtherosclerosisDocument5 pagesEngineering Grafts: Genetic Vein Resistant AtherosclerosisTheo KhouryNo ratings yet
- Acute Kidney Injury Pathology and PathophysiologyDocument11 pagesAcute Kidney Injury Pathology and Pathophysiologyhossein kasiriNo ratings yet
- Rocha Singh2014Document9 pagesRocha Singh2014Riandhani UpitariniaNo ratings yet
- Patten 2000Document6 pagesPatten 2000diqa aridaniNo ratings yet
- Hepatic Stellate Cells and Liver FibrosisDocument20 pagesHepatic Stellate Cells and Liver Fibrosiscc vereNo ratings yet
- Nutrition and AtherosclerosisDocument19 pagesNutrition and AtherosclerosisJOSE RICARDO LOPEZ PALESTINANo ratings yet
- 1 s2.0 S2542568420300386 MainDocument7 pages1 s2.0 S2542568420300386 MainLaila Kurnia PramonoNo ratings yet
- Nihms 260877Document10 pagesNihms 260877febyan yohanesNo ratings yet
- Trombo y Aaa PDFDocument11 pagesTrombo y Aaa PDFAnna Sofía ParedesNo ratings yet
- Jurnal PendarahanDocument15 pagesJurnal Pendarahanイアン リムホト ザナガNo ratings yet
- Overview of and Approach To The Vasculitides in Adults - UpToDateDocument25 pagesOverview of and Approach To The Vasculitides in Adults - UpToDateArthurNo ratings yet
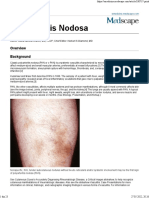
- Poliarterita NodoasăDocument23 pagesPoliarterita NodoasăElena StepanNo ratings yet
- Calcification in Atherosclerotic Plaque Vulnerability - Friend or Foe - 2020Document12 pagesCalcification in Atherosclerotic Plaque Vulnerability - Friend or Foe - 2020Olry Le NobelNo ratings yet
- Sfaa 142Document11 pagesSfaa 142Eduardo AdameNo ratings yet
- Pathogenesis of Atherosclerosis A Review PDFDocument7 pagesPathogenesis of Atherosclerosis A Review PDFPrecious IJNo ratings yet
- 1 s2.0 S141335552300014X MainDocument11 pages1 s2.0 S141335552300014X MainFirmansyah PurwantoNo ratings yet
- Left Atrial Appendage Occlusion 1Document37 pagesLeft Atrial Appendage Occlusion 1Antonia AdinaNo ratings yet
- AtvbahaDocument16 pagesAtvbahaapi-654112138No ratings yet
- GFQ 356Document8 pagesGFQ 356Ricardo IbarraNo ratings yet
- Review Hyperglycemia and Endothelial Dysfunction in AtherosclerosisDocument19 pagesReview Hyperglycemia and Endothelial Dysfunction in AtherosclerosisTakdirtahirNo ratings yet
- An Anatomical and Radiological Study of Origins of The Arteries Forming The Celiac Trunk: Clinical and Embryological ImplicationsDocument15 pagesAn Anatomical and Radiological Study of Origins of The Arteries Forming The Celiac Trunk: Clinical and Embryological ImplicationsTung MoNo ratings yet
- Study PCOS ObesityDocument9 pagesStudy PCOS ObesitydfdfffffNo ratings yet
- Bicuspid AorticDocument12 pagesBicuspid Aorticikbal rambalinoNo ratings yet
- 1 s2.0 S2666602223000125 MainDocument6 pages1 s2.0 S2666602223000125 Mainosa345484No ratings yet
- Assessment of AtherosclerosisDocument10 pagesAssessment of AtherosclerosisCalysta CitraNo ratings yet
- Uremic BleedingDocument16 pagesUremic BleedingSofia SimpertigueNo ratings yet
- Overview of and Approach To The Vasculitides in Adults - UpToDateDocument26 pagesOverview of and Approach To The Vasculitides in Adults - UpToDateElena StepanNo ratings yet
- Epoetin Treatment: What Are The Arguments To Expect A Beneficial Effect On Renal Disease Progression?Document4 pagesEpoetin Treatment: What Are The Arguments To Expect A Beneficial Effect On Renal Disease Progression?Mohamad IrwanNo ratings yet
- Fistula ArteriovenosaDocument14 pagesFistula ArteriovenosaJoshua CajinaNo ratings yet
- Treatment Strategies of NSTEMI-ACS With Multivessel Disease: Gani Bajraktari, Michael Y. HeneinDocument3 pagesTreatment Strategies of NSTEMI-ACS With Multivessel Disease: Gani Bajraktari, Michael Y. HeneinolisNo ratings yet
- The Role of Basement Membranes in Cerebral Amyloid AngiopathyDocument20 pagesThe Role of Basement Membranes in Cerebral Amyloid AngiopathyAndrade GuiNo ratings yet
- WJH 13 533Document11 pagesWJH 13 533CIRUGÍA ONCOLÓGICA ABDOMENNo ratings yet
- AtherosclerosisDocument38 pagesAtherosclerosisaffansungkarNo ratings yet
- Early Atherosclerotic Changes in Coronary Arteries Are Associated With Endothelium Shear Stress Contraction Expansion VariabilityDocument16 pagesEarly Atherosclerotic Changes in Coronary Arteries Are Associated With Endothelium Shear Stress Contraction Expansion Variabilityhassenesalam18No ratings yet
- Chatterjee 2017Document21 pagesChatterjee 2017Yohana SetiawanNo ratings yet
- NETs Complicaciones Transplante Higado Fimmu-13-1054753Document14 pagesNETs Complicaciones Transplante Higado Fimmu-13-1054753Silvana MaldonadoNo ratings yet
- Japplphysiol 00035 2004Document10 pagesJapplphysiol 00035 2004Kritik KumarNo ratings yet
- Dagher, Therapeutic Potential of Quercetin To Alleviate Endothelial Dysfunction in Age-Related Cardiovascular Diseases ODocument24 pagesDagher, Therapeutic Potential of Quercetin To Alleviate Endothelial Dysfunction in Age-Related Cardiovascular Diseases ODaniel Castillo hernándezNo ratings yet
- Overview of and Approach To The Vasculitides in AdultsDocument25 pagesOverview of and Approach To The Vasculitides in AdultsHello AngelNo ratings yet
- Pathophysiology and Mechanisms of Acute Coronary SyndromesDocument13 pagesPathophysiology and Mechanisms of Acute Coronary SyndromesShawnNo ratings yet
- Microcirculation in Cardiovascular DiseasesFrom EverandMicrocirculation in Cardiovascular DiseasesEnrico Agabiti-RoseiNo ratings yet
- Tarea 2Document18 pagesTarea 2Verónica SolísNo ratings yet
- Ajc 16 12 980 989Document10 pagesAjc 16 12 980 989Maria Goretty L. TobingNo ratings yet
- 2018 Cartilage Regeneration in Humans With Adipose Tissue-Derived Stem Cells and Adipose Stromal Vascular Fraction CellsDocument18 pages2018 Cartilage Regeneration in Humans With Adipose Tissue-Derived Stem Cells and Adipose Stromal Vascular Fraction Cells林煒No ratings yet
- Perivascular Adipose Tissue 2023 MDocument24 pagesPerivascular Adipose Tissue 2023 Mmiguel contrerasNo ratings yet
- Perivascular Adipose Tissue 2023 QDocument14 pagesPerivascular Adipose Tissue 2023 Qmiguel contrerasNo ratings yet
- Perivascular Adipose Tissue 2023 BDocument18 pagesPerivascular Adipose Tissue 2023 Bmiguel contrerasNo ratings yet
- Perivascular Adipose Tissue 2023 IDocument9 pagesPerivascular Adipose Tissue 2023 Imiguel contrerasNo ratings yet
- Cardio Diabetes 1Document15 pagesCardio Diabetes 1miguel contrerasNo ratings yet
- Guias Hipertension 2023 EscDocument199 pagesGuias Hipertension 2023 Escmiguel contrerasNo ratings yet
- SGLT2 Ahf2Document10 pagesSGLT2 Ahf2miguel contrerasNo ratings yet
- The Biological Perspective 1Document18 pagesThe Biological Perspective 1Radhey SurveNo ratings yet
- CA Stomach OverviewDocument84 pagesCA Stomach OverviewDr Shafiq Ahmad ChughtaiNo ratings yet
- Macroscopic Diagnostic For Pathoanathomy and Cytopathology ExamDocument21 pagesMacroscopic Diagnostic For Pathoanathomy and Cytopathology Examkapil pancholiNo ratings yet
- Macrophage and Osteoblast Response To Micro and Nano Hydroxyapatite - A ReviewDocument13 pagesMacrophage and Osteoblast Response To Micro and Nano Hydroxyapatite - A Reviewnano_journalNo ratings yet
- Atrophic Pap Smears, Differential Diagnosis and Pitfalls: A ReviewDocument5 pagesAtrophic Pap Smears, Differential Diagnosis and Pitfalls: A ReviewJill ValdesNo ratings yet
- Autonomic Nervous SystemDocument28 pagesAutonomic Nervous Systemglad mohesh100% (1)
- Endocrine Lab ManualDocument9 pagesEndocrine Lab ManualApril Joy de LimaNo ratings yet
- The Seven Chakras of Your Body and Their SignificanceDocument4 pagesThe Seven Chakras of Your Body and Their SignificancerudraarjunNo ratings yet
- Disseminated Intravascular CoagulationDocument14 pagesDisseminated Intravascular CoagulationAmanda ScarletNo ratings yet
- Tissues: 1. Objective QuestionsDocument7 pagesTissues: 1. Objective Questionsnaman mahawer100% (1)
- Package Insert - DG Gel 8 ABO-Rh (2D)Document10 pagesPackage Insert - DG Gel 8 ABO-Rh (2D)Liliana MirandaNo ratings yet
- NMAT Biology Practice Questions Set 1Document6 pagesNMAT Biology Practice Questions Set 1Nurshayma JalilNo ratings yet
- ImmunologyDocument1 pageImmunologyChris_Barber0986% (7)
- Skeletal System Important Qs Important Questions ICSE 995963bf 28ef 48c2 Ad56 5a8b29c84771Document2 pagesSkeletal System Important Qs Important Questions ICSE 995963bf 28ef 48c2 Ad56 5a8b29c84771sujata duttaNo ratings yet
- Spine AnatomyDocument8 pagesSpine Anatomyvishramki2009No ratings yet
- Detailed Lesson Plan in Anatomy and PhysiologyDocument7 pagesDetailed Lesson Plan in Anatomy and PhysiologyMea DolautaNo ratings yet
- Esophageal Carcinoma: Preceptor: Dr. Ratesh KhillanDocument37 pagesEsophageal Carcinoma: Preceptor: Dr. Ratesh KhillanVarun VarnNo ratings yet
- The Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionDocument14 pagesThe Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionPaolaNo ratings yet
- Genitourinary AssessementDocument46 pagesGenitourinary AssessementShenaNo ratings yet
- DR - Nugroho Tjandra, Sp. Rad Staf Fungsiomal Radiologi Rumkital DR Ramelan Pengajar FK UHTDocument49 pagesDR - Nugroho Tjandra, Sp. Rad Staf Fungsiomal Radiologi Rumkital DR Ramelan Pengajar FK UHTeddo.leonardoNo ratings yet
- Structure and Function of CellDocument11 pagesStructure and Function of CellWoobin RealNo ratings yet
- Human Reproduction DPP-03Document3 pagesHuman Reproduction DPP-03vijaylakshmi0727No ratings yet
- Bio 11 Lab Exam 3 ReviewerDocument5 pagesBio 11 Lab Exam 3 Revieweririsiri28No ratings yet
- Head and Neck Who ClassificationDocument3 pagesHead and Neck Who ClassificationLorena Paula0% (1)
- Functional Ultrastructure An Atlas of Tissue Biology and PathologyDocument341 pagesFunctional Ultrastructure An Atlas of Tissue Biology and PathologyАндрей СкрынникNo ratings yet
- Skeletal System Lab ReportDocument7 pagesSkeletal System Lab Reportapi-296603992No ratings yet
- Waec Biology ExampulseDocument82 pagesWaec Biology ExampulsealbertayomideakiodeNo ratings yet
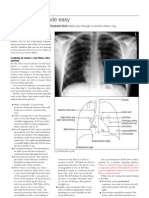
- Cara Membaca Foto Thoraks Yang BaikDocument2 pagesCara Membaca Foto Thoraks Yang BaikIdi Nagan RayaNo ratings yet
- Lab 6 - Overview of The Skeletal SystemDocument6 pagesLab 6 - Overview of The Skeletal Systemsidro123No ratings yet
- B.ing YenishacDocument3 pagesB.ing YenishacYenisha CherinaNo ratings yet