You might also like
- Research Article Curcumin Attenuates Ferroptosis and Ameliorates Erectile Function in Diabetic Rats by Activating Nrf2/HO-1 PathwayDocument13 pagesResearch Article Curcumin Attenuates Ferroptosis and Ameliorates Erectile Function in Diabetic Rats by Activating Nrf2/HO-1 Pathwaymohammed fayedNo ratings yet
- Correos Electrónicos Active Vitamin D3 Protects Against Diabetic Kidney Disease by Regulating The JNK Signaling Pathway in RatsDocument9 pagesCorreos Electrónicos Active Vitamin D3 Protects Against Diabetic Kidney Disease by Regulating The JNK Signaling Pathway in Ratsnoranton7No ratings yet
- Ciglitazone, A PPARy Agonist, Ameliorates Diabetic Nephropaty in Part Through Homocysteine ClearanceDocument9 pagesCiglitazone, A PPARy Agonist, Ameliorates Diabetic Nephropaty in Part Through Homocysteine ClearanceBrian KuslerNo ratings yet
- Review Article: Streptozotocin-Induced Diabetes Models: Pathophysiological Mechanisms and Fetal OutcomesDocument12 pagesReview Article: Streptozotocin-Induced Diabetes Models: Pathophysiological Mechanisms and Fetal OutcomesMayra PereiraNo ratings yet
- Wang 2013Document7 pagesWang 2013Archisman MahapatraNo ratings yet
- PLD - Systematic ReviewDocument23 pagesPLD - Systematic ReviewCarleta StanNo ratings yet
- Transcriptomic and Proteomic Analysis of Potential Therapeutic Target Genes in The Liver of Metformin Treated Sprague Dawley Rats With Type 2 Diabetes Mellitusijmm-41!06!3327Document15 pagesTranscriptomic and Proteomic Analysis of Potential Therapeutic Target Genes in The Liver of Metformin Treated Sprague Dawley Rats With Type 2 Diabetes Mellitusijmm-41!06!3327ErickNo ratings yet
- Enmet DM AntioxDocument9 pagesEnmet DM AntioxAreef MuarifNo ratings yet
- Intermittent Hypoxia-Activated Cyclooxygenase Pathway: Role in AtherosclerosisDocument10 pagesIntermittent Hypoxia-Activated Cyclooxygenase Pathway: Role in AtherosclerosisaditNo ratings yet
- ... Bao Et Al2015Document8 pages... Bao Et Al2015Gabriel GursenNo ratings yet
- Ijms 22 05727Document12 pagesIjms 22 05727nur chasanahNo ratings yet
- Garlic (Allium Sativum) and Ginger (Zingiber Officinale) Attenuate Structural Nephropathy Progression in Streptozotocin-Induced Diabetic RatsDocument10 pagesGarlic (Allium Sativum) and Ginger (Zingiber Officinale) Attenuate Structural Nephropathy Progression in Streptozotocin-Induced Diabetic RatsAna HerawatiNo ratings yet
- Ijcmr 2510 PDFDocument5 pagesIjcmr 2510 PDFAtul DwivediNo ratings yet
- Quercetin PankreasDocument7 pagesQuercetin PankreasAlmas TNo ratings yet
- Swietenine NAFLDDocument15 pagesSwietenine NAFLDpmallirao2000No ratings yet
- HHS Public Access: Highlighting Diabetes - The Epidemic ContinuesDocument14 pagesHHS Public Access: Highlighting Diabetes - The Epidemic ContinuesRidhaNo ratings yet
- Thesis Proposal.Document27 pagesThesis Proposal.Edrick RamoranNo ratings yet
- Increased Islet Apoptosis in Pdx1 MiceDocument14 pagesIncreased Islet Apoptosis in Pdx1 Miceroh009No ratings yet
- 5570796Document9 pages5570796Leroy Christy LawalataNo ratings yet
- BMC Clinical Pathology: Protection of Early Phase Hepatic Ischemia-Reperfusion Injury by Cholinergic AgonistsDocument13 pagesBMC Clinical Pathology: Protection of Early Phase Hepatic Ischemia-Reperfusion Injury by Cholinergic AgonistsPrachi KariaNo ratings yet
- KersenDocument13 pagesKersenJumasni AdnanNo ratings yet
- Gale Et Al 2011 Disruption of Circadian Rhythms Accelerates Development of Diabetes Through Pancreatic Beta Cell LossDocument11 pagesGale Et Al 2011 Disruption of Circadian Rhythms Accelerates Development of Diabetes Through Pancreatic Beta Cell Lossleila chebahNo ratings yet
- Effect of Melatonin and Chromium Picolinate Administration To Highcarbohydrate Dietfed Male Wistar Rats 1747 0862 1000245Document7 pagesEffect of Melatonin and Chromium Picolinate Administration To Highcarbohydrate Dietfed Male Wistar Rats 1747 0862 1000245jovaneticNo ratings yet
- Circadian clocks linked to diabetes and heart diseaseDocument8 pagesCircadian clocks linked to diabetes and heart diseaseSakshi JoshiNo ratings yet
- Metformin Attenuates Early-Stage Atherosclerosis in Mildly Hyperglycemic Oikawa-Nagao MiceDocument9 pagesMetformin Attenuates Early-Stage Atherosclerosis in Mildly Hyperglycemic Oikawa-Nagao Micegandik danu pramigaNo ratings yet
- Noninvasive Magnetic Resonance Imaging of Microvascular Changes in Type 1 DiabetesDocument6 pagesNoninvasive Magnetic Resonance Imaging of Microvascular Changes in Type 1 DiabetesAndrés Esteban Quezada VenegasNo ratings yet
- Bmri2018 4706976Document11 pagesBmri2018 4706976Denny BunNo ratings yet
- Therapeutic Potential of Nrf2 Activators in Streptozotocin-Induced Diabetic NephropathyDocument12 pagesTherapeutic Potential of Nrf2 Activators in Streptozotocin-Induced Diabetic NephropathyAgustinus VincentNo ratings yet
- Research Article: Recovery of The Cell Cycle Inhibition in CCL - Induced Cirrhosis by The Adenosine Derivative Ifc-305Document13 pagesResearch Article: Recovery of The Cell Cycle Inhibition in CCL - Induced Cirrhosis by The Adenosine Derivative Ifc-305Indri Noor HidayatiNo ratings yet
- Continuous Erythropoietin Receptor Activator Ameliorates Diabetic Kidney Disease in RatsDocument9 pagesContinuous Erythropoietin Receptor Activator Ameliorates Diabetic Kidney Disease in RatsAli SahinNo ratings yet
- Metformin protects hepatocytes from oxidative stressDocument12 pagesMetformin protects hepatocytes from oxidative stressyasmin matterNo ratings yet
- Ketogenic Diet Benefits in DiabetesDocument9 pagesKetogenic Diet Benefits in DiabetesMitchel Córdova LoaizaNo ratings yet
- IL-2 and IFN-gamma in The Retina of Diabetic Rats: Basic ScienceDocument7 pagesIL-2 and IFN-gamma in The Retina of Diabetic Rats: Basic ScienceRichard UldebrandoNo ratings yet
- Empagliflozin Is Not Renoprotective in Non-Diabetic Rat Models of Chronic Kidney DiseaseDocument14 pagesEmpagliflozin Is Not Renoprotective in Non-Diabetic Rat Models of Chronic Kidney DiseasesamuelNo ratings yet
- Diabetes Mellitus Directs NKT Cells Toward Type 2 and Regulatory PhenotypeDocument7 pagesDiabetes Mellitus Directs NKT Cells Toward Type 2 and Regulatory PhenotypefragaNo ratings yet
- The Cellular Fate of Glucose and Its Relevance In-1-16Document16 pagesThe Cellular Fate of Glucose and Its Relevance In-1-16Yariela Amador MendietaNo ratings yet
- Sunday O. Oyedemi - Godswill Nwaogu - Chika I. Chukwuma - Olaoluwa T. Adeyemi - Motlalepula G. Matsabisa - Shasank S. Swain - Olayinka A. AiyegoroDocument16 pagesSunday O. Oyedemi - Godswill Nwaogu - Chika I. Chukwuma - Olaoluwa T. Adeyemi - Motlalepula G. Matsabisa - Shasank S. Swain - Olayinka A. AiyegoroGiovanni OliveiraNo ratings yet
- Effect of Mesenchymal Stem Cells Transplantation On Glycaemic Profile & Their Localization in Streptozotocin Induced Diabetic Wistar RatsDocument9 pagesEffect of Mesenchymal Stem Cells Transplantation On Glycaemic Profile & Their Localization in Streptozotocin Induced Diabetic Wistar RatsShanaz NovriandinaNo ratings yet
- 2017 Article 199Document13 pages2017 Article 199Rabiatul 'raney' AdawiyahNo ratings yet
- Evaluation of Fresh Frozen Plasma Administration in Dogs With Pancreatitis: 77 Cases (1995 2005)Document6 pagesEvaluation of Fresh Frozen Plasma Administration in Dogs With Pancreatitis: 77 Cases (1995 2005)soledadDC329No ratings yet
- The Essential Role of Endothelial Nitric Oxide Synthase Activation in Insulin-Mediated Neuroprotection Against Ischemic Stroke in DiabetesDocument9 pagesThe Essential Role of Endothelial Nitric Oxide Synthase Activation in Insulin-Mediated Neuroprotection Against Ischemic Stroke in DiabetesrivandiNo ratings yet
- Cell Biochem Funct 2011 p55Document9 pagesCell Biochem Funct 2011 p55IQBAL HAKKIKINo ratings yet
- Nihms 989444Document22 pagesNihms 989444Guillermo Angel Herrera ChavezNo ratings yet
- Research Article Clostridium Butyricum AgainstDocument13 pagesResearch Article Clostridium Butyricum Againstos krishnaNo ratings yet
- Fang 2005Document10 pagesFang 2005pamela gaitanNo ratings yet
- Review Article: Acupoint Therapy On Diabetes Mellitus and Its Common Chronic Complications: A Review of Its MechanismsDocument10 pagesReview Article: Acupoint Therapy On Diabetes Mellitus and Its Common Chronic Complications: A Review of Its Mechanismsoscarin123456789No ratings yet
- Insulin Stimulates Mitochondrial FusionDocument14 pagesInsulin Stimulates Mitochondrial Fusionmercuryboss90No ratings yet
- Vaithilingam Review RDS-2017 3Document28 pagesVaithilingam Review RDS-2017 3Voltaire Méndez RodríguezNo ratings yet
- Artificially Cultivated Ophiocordyceps Sinensis AlleviatesDocument17 pagesArtificially Cultivated Ophiocordyceps Sinensis AlleviatesHoang IceNo ratings yet
- Reactive Oxygen and Nitrogen Species in Sepsis-Induced Hepatic Microvascular DysfunctionDocument10 pagesReactive Oxygen and Nitrogen Species in Sepsis-Induced Hepatic Microvascular DysfunctionGiosué Reyes HernandezNo ratings yet
- GLP AlzhmrDocument14 pagesGLP Alzhmryengeç adamNo ratings yet
- Gene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GeneDocument7 pagesGene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GenesaifudinNo ratings yet
- Rodent Models of Streptozotocin-Induced Diabetic NephropathyDocument6 pagesRodent Models of Streptozotocin-Induced Diabetic NephropathyManishankar PampanaNo ratings yet
- Journal of Translational MedicineDocument13 pagesJournal of Translational MedicinetrirahmiNo ratings yet
- Animals 13 00684Document18 pagesAnimals 13 00684erni manikNo ratings yet
- 5.ashim R2Document12 pages5.ashim R2Asim DuttaNo ratings yet
- Economic Pigging Cycles For Low-Throughput PipelinesDocument11 pagesEconomic Pigging Cycles For Low-Throughput Pipelineseng.Abdelaziz RedaNo ratings yet
- Vol-1, Issue 7 Paper (5) Page 26-31Document7 pagesVol-1, Issue 7 Paper (5) Page 26-31IJMSHCNo ratings yet
- Type 2 Diabetes Pathogenesis: Insulin Resistance and Beta Cell DysfunctionDocument324 pagesType 2 Diabetes Pathogenesis: Insulin Resistance and Beta Cell DysfunctionJesus Eduardo Cruz-BurgoinNo ratings yet
- Time TableDocument28 pagesTime TablePetWise Animal ClinicNo ratings yet
- HRSL ScriptsDocument3 pagesHRSL ScriptsFolajimi AdebowaleNo ratings yet
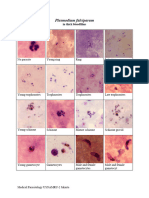
- Digital Picture Human MalariaDocument8 pagesDigital Picture Human MalariaOlivia ChricilloaNo ratings yet
- New Nursing Diagnoses 2012-2014Document11 pagesNew Nursing Diagnoses 2012-2014CarmenNoemiSantos100% (1)
- 1 Module 1 hss1010 Booklet 1Document108 pages1 Module 1 hss1010 Booklet 1api-463761938No ratings yet
- Critical Values List: Arterial Blood GasesDocument3 pagesCritical Values List: Arterial Blood Gasesiq_dianaNo ratings yet
- Palm Grass Blood Clot StudyDocument20 pagesPalm Grass Blood Clot StudyRichele QuilantangNo ratings yet
- Clinical Pathology - Peripheral Blood Smear - Part 1 (Dr. Carretas)Document7 pagesClinical Pathology - Peripheral Blood Smear - Part 1 (Dr. Carretas)ashley nicholeNo ratings yet
- Poisons of Plant OriginDocument46 pagesPoisons of Plant OriginThe AbyssinicansNo ratings yet
- 12 MRCP 2 Opthalmology NOTES PassmedicineDocument30 pages12 MRCP 2 Opthalmology NOTES PassmedicineHussein GhannamNo ratings yet
- Introduction to Virology Lec.1Document20 pagesIntroduction to Virology Lec.1Umer ZohaibNo ratings yet
- Micro-Chem Plus TDS English GHSDocument1 pageMicro-Chem Plus TDS English GHSFrank AlvaradoNo ratings yet
- Malaria Updates in Indonesia - KemenkesDocument17 pagesMalaria Updates in Indonesia - KemenkesAmirullah ArsyadNo ratings yet
- Skin CancerDocument24 pagesSkin CancerMister David anthonyNo ratings yet
- Reduce cholesterol and triglycerides with CRESTORDocument2 pagesReduce cholesterol and triglycerides with CRESTORSunny Mae T. PuigNo ratings yet
- CASE STUDY (Gastro)Document3 pagesCASE STUDY (Gastro)Jake Yvan DizonNo ratings yet
- Pharmaceutical Care PlaneDocument28 pagesPharmaceutical Care PlaneMUGHALNo ratings yet
- SCIENCEDocument6 pagesSCIENCEFerlyn SolimaNo ratings yet
- Disturbed Sensory PerceptionDocument3 pagesDisturbed Sensory PerceptionJashtine JingcoNo ratings yet
- Presentation Advance Nursing PracticeDocument33 pagesPresentation Advance Nursing PracticeSandesh LokhandeNo ratings yet
- Leukemia in Children: 1 Rahul Dhaker, Asst. Professor, RCNDocument41 pagesLeukemia in Children: 1 Rahul Dhaker, Asst. Professor, RCNRahul DhakerNo ratings yet
- Oseltamivir Indication, Dosage, Side Effect, Precaution MIMS IndonesiaDocument1 pageOseltamivir Indication, Dosage, Side Effect, Precaution MIMS Indonesiaintan alvinnNo ratings yet
- Proposal Form: Flexi HealthDocument5 pagesProposal Form: Flexi HealthSANDEEP familyNo ratings yet
- DRRR Module 4 5Document4 pagesDRRR Module 4 5kellan lyfeNo ratings yet
- Internal DiseaseDocument55 pagesInternal Diseaseshilpa sekhar278No ratings yet
- Fip Call To Action: To Support Pharmacists and Pharmacy Workers On The coronavirus/COVID-19 FrontlineDocument7 pagesFip Call To Action: To Support Pharmacists and Pharmacy Workers On The coronavirus/COVID-19 Frontlinebenny christantoNo ratings yet
- Yoga Mudras For Balancing Dosha (Vata, Pitta, & Kapha) : Home Articles Mudras Poses PranayamaDocument26 pagesYoga Mudras For Balancing Dosha (Vata, Pitta, & Kapha) : Home Articles Mudras Poses PranayamaDanaNo ratings yet
- Brain - Drain The Impact of Air Pulution On Firm PerformanceDocument69 pagesBrain - Drain The Impact of Air Pulution On Firm PerformancesinhNo ratings yet
- The Aspects of GENE THERAPYDocument12 pagesThe Aspects of GENE THERAPYJasmin Prindiana100% (6)
- Passed Like A ShadowDocument15 pagesPassed Like A ShadowTimoNo ratings yet