You might also like
- Exploring The Neural ManifoldDocument4 pagesExploring The Neural ManifoldKarla ValderramaNo ratings yet
- Context-Dependent Control of Behavior in Drosophila PDFDocument9 pagesContext-Dependent Control of Behavior in Drosophila PDFMert GencNo ratings yet
- The Brain from Inside Out Chapter 4 SummaryDocument20 pagesThe Brain from Inside Out Chapter 4 SummaryjuannnnNo ratings yet
- Large-Scale Neural Recordings Call For New Insights To Link Brain and BehaviorDocument9 pagesLarge-Scale Neural Recordings Call For New Insights To Link Brain and BehaviorMoshe AlonsoNo ratings yet
- Rizzolatti 1998 Language PDFDocument7 pagesRizzolatti 1998 Language PDFappled.apNo ratings yet
- Criteria For Consciousness in Humans and Other Mammals: Anil K. Seth, Bernard J. Baars, David B. EdelmanDocument21 pagesCriteria For Consciousness in Humans and Other Mammals: Anil K. Seth, Bernard J. Baars, David B. Edelmankfun90No ratings yet
- Communication From The Cerebellum To The Neocortex During Sleep SpindlesDocument1 pageCommunication From The Cerebellum To The Neocortex During Sleep SpindlesDavid NazarovNo ratings yet
- Is Neuroimaging Measuring Information in The BrainDocument15 pagesIs Neuroimaging Measuring Information in The BrainAndré KimNo ratings yet
- Sensation During Active BehaviorsDocument9 pagesSensation During Active BehaviorsJuliana Ocampo FernandezNo ratings yet
- 1 s2.0 S089662731931044X MainDocument19 pages1 s2.0 S089662731931044X MainJorge Miranda JaimeNo ratings yet
- Resting State fMRI Distinguishes Human Ventral Tegmental Area and Substantia NigraDocument24 pagesResting State fMRI Distinguishes Human Ventral Tegmental Area and Substantia NigraErick SolisNo ratings yet
- The Steady-State Visual Evoked Potential in Vision Research A ReviewDocument46 pagesThe Steady-State Visual Evoked Potential in Vision Research A Review6089272No ratings yet
- Linkage Disequilibrium in The Human GenomeDocument7 pagesLinkage Disequilibrium in The Human GenomedioufNo ratings yet
- Cerebro de MoscaDocument37 pagesCerebro de Moscafernando gonzalezNo ratings yet
- Neuronal Representation of Saccadic Error in Macaque Posterior Parietal Cortex (PPC)Document17 pagesNeuronal Representation of Saccadic Error in Macaque Posterior Parietal Cortex (PPC)Zeus HeraklesNo ratings yet
- The Neural Code For Motor Control in The Cerebellum and Oculomotor BrainstemDocument16 pagesThe Neural Code For Motor Control in The Cerebellum and Oculomotor BrainstemPrivat EtavirpNo ratings yet
- Perspective: Neuronal Cell Types and Connectivity: Lessons From The RetinaDocument11 pagesPerspective: Neuronal Cell Types and Connectivity: Lessons From The Retinaports1111No ratings yet
- CSCS24 Paper 42 yDocument6 pagesCSCS24 Paper 42 yDaniela HossuNo ratings yet
- Elife 61881 v3Document26 pagesElife 61881 v3Pegah KassraianNo ratings yet
- Neural Plasticity and Consciousness (Figures Are Missing in This Version) Susan HurleyDocument35 pagesNeural Plasticity and Consciousness (Figures Are Missing in This Version) Susan HurleyA_WretchNo ratings yet
- Light affects mood and learning through distinct retina-brain pathwaysDocument33 pagesLight affects mood and learning through distinct retina-brain pathwaysJaime RoldanNo ratings yet
- Choosing The Greater of Two Goods: Neural Currencies For Valuation and Decision MakingDocument14 pagesChoosing The Greater of Two Goods: Neural Currencies For Valuation and Decision MakingMichel Andres Patiño SaenzNo ratings yet
- Buzsaki 2004 Nat NeurosciDocument7 pagesBuzsaki 2004 Nat Neuroscihussin elrashidyNo ratings yet
- Vision Research: David H. Mcdougal, Paul D. GamlinDocument16 pagesVision Research: David H. Mcdougal, Paul D. GamlinCristina GómezNo ratings yet
- Reconstruction of Virtual Neural Circuits in An Insect BrainDocument8 pagesReconstruction of Virtual Neural Circuits in An Insect BrainnithiananthiNo ratings yet
- Neuronas de La RetinaDocument10 pagesNeuronas de La Retinaxime20031410No ratings yet
- A Dual Role For Prediction Error inDocument11 pagesA Dual Role For Prediction Error inmiguel.amendozaNo ratings yet
- Nocturnal Light Exposure Impairs Affective Responses in A Wavelength-Dependent MannerDocument7 pagesNocturnal Light Exposure Impairs Affective Responses in A Wavelength-Dependent MannerLiana DimaNo ratings yet
- NOISE GoldmanPLOS06Document8 pagesNOISE GoldmanPLOS06Nguyen HienNo ratings yet
- Context-Specific Habituation A ReviewDocument11 pagesContext-Specific Habituation A Reviewomar.a.moslehNo ratings yet
- TMP 5 FADocument12 pagesTMP 5 FAFrontiersNo ratings yet
- Neural Manifold Analysis of Brain Circuit DynamicsDocument24 pagesNeural Manifold Analysis of Brain Circuit DynamicsKarla ValderramaNo ratings yet
- Neuro PDFDocument4 pagesNeuro PDFChris MacknightNo ratings yet
- Dynamic BrainDocument35 pagesDynamic BrainchandusgNo ratings yet
- An Extreme Value Theory Model of Cross-Modal Sensory Information Integration in Modulation of Vertebrate Visual System FunctionsDocument18 pagesAn Extreme Value Theory Model of Cross-Modal Sensory Information Integration in Modulation of Vertebrate Visual System Functionswill bNo ratings yet
- The Steady-State Visual Evoked Potential in Vision Research: A ReviewDocument46 pagesThe Steady-State Visual Evoked Potential in Vision Research: A Reviewkickinghorse892No ratings yet
- Gollisch MeisterDocument15 pagesGollisch MeisterElaheh LotfiNo ratings yet
- 10 1126@science Aau8302Document18 pages10 1126@science Aau8302nicolasNo ratings yet
- Sistemul AuditivDocument10 pagesSistemul AuditivCalugareanu MadalinaNo ratings yet
- 174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperienceDocument20 pages174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperiencejuannnnNo ratings yet
- Synapsigvdrts en SueñoDocument11 pagesSynapsigvdrts en SueñoLEISLY TATIANA OVIEDO GOMEZNo ratings yet
- Brain Rhythms and Neural Syntax Implications For Efficient Coding of Cognitive Content and Neuropsychiatric DiseaseDocument24 pagesBrain Rhythms and Neural Syntax Implications For Efficient Coding of Cognitive Content and Neuropsychiatric Diseaselionita sugandiNo ratings yet
- 1Document11 pages1Almar ZamNo ratings yet
- Perceptual Learning and Adult Cortical Plasticity2009Journal of PhysiologyDocument9 pagesPerceptual Learning and Adult Cortical Plasticity2009Journal of Physiologyshefaligupta91No ratings yet
- Towards An Empirically Grounded Predictive Coding Account of Action UnderstandingDocument3 pagesTowards An Empirically Grounded Predictive Coding Account of Action UnderstandingAlan HartogNo ratings yet
- 1 s2.0 S096098221730708X MainDocument5 pages1 s2.0 S096098221730708X Mainrotinda bilekNo ratings yet
- Neural Encoding and Decoding with Deep LearningDocument27 pagesNeural Encoding and Decoding with Deep LearningUsman KhalidNo ratings yet
- Overview CerebellumDocument21 pagesOverview CerebellumwilsonnianNo ratings yet
- 2.2 Encoding - InformationDocument15 pages2.2 Encoding - InformationYujie WangNo ratings yet
- TMP 64 B7Document18 pagesTMP 64 B7FrontiersNo ratings yet
- Curing TB With SunlightDocument1 pageCuring TB With SunlightLawrence NapitupuluNo ratings yet
- Deprivation-Induced Plasticity in The Early Central Circuits of The Rodent Visual, Auditory, and Olfactory Systems 2024Document21 pagesDeprivation-Induced Plasticity in The Early Central Circuits of The Rodent Visual, Auditory, and Olfactory Systems 2024anacarolinadepaduaaNo ratings yet
- NPH 011 033403Document28 pagesNPH 011 033403Cristhian RodriguezNo ratings yet
- Measuring Shared Responses Across Subjects Using Intersubject CorrelationDocument36 pagesMeasuring Shared Responses Across Subjects Using Intersubject CorrelationAnonymous HUY0yRexYfNo ratings yet
- Synapse and MemoryDocument19 pagesSynapse and Memorysomya mathurNo ratings yet
- Biopsych 2Document7 pagesBiopsych 2Arshia RoyNo ratings yet
- Download Generating Artificial Sensations With Spinal Cord Stimulation In Primates And Rodents Amol P Yadav Shuangyan Li Max O Krucoff Mikhail A Lebedev Muhammad M Abd El Barr Miguel A L Nicolel full chapterDocument35 pagesDownload Generating Artificial Sensations With Spinal Cord Stimulation In Primates And Rodents Amol P Yadav Shuangyan Li Max O Krucoff Mikhail A Lebedev Muhammad M Abd El Barr Miguel A L Nicolel full chapterelmer.coyle542100% (3)
- TMP 286 ADocument14 pagesTMP 286 AFrontiersNo ratings yet
- Mitra Et Al 2016 How Networks Communicate Propagation Patterns in Spontaneous Brain ActivityDocument9 pagesMitra Et Al 2016 How Networks Communicate Propagation Patterns in Spontaneous Brain ActivityVasillis MamosNo ratings yet
- Engine Technology Course A/F Control Classroom TrainingDocument14 pagesEngine Technology Course A/F Control Classroom Traininglongtrandang5867No ratings yet
- Gripper of FTV Invisio Panels For Horizontal FacadeDocument16 pagesGripper of FTV Invisio Panels For Horizontal FacadefghfNo ratings yet
- SRT95 Off-Highway Truck Rear Axle Assembly: Structure and Installation Part 2 Parts Presentation Part 3 MaintenanceDocument29 pagesSRT95 Off-Highway Truck Rear Axle Assembly: Structure and Installation Part 2 Parts Presentation Part 3 MaintenanceoktopusNo ratings yet
- (World of Art) John Boardman - Athenian Black Figure Vases-Thames and Hudson (1974)Document128 pages(World of Art) John Boardman - Athenian Black Figure Vases-Thames and Hudson (1974)Karen SantosNo ratings yet
- OTISLINE QuestionsDocument5 pagesOTISLINE QuestionsArvind Gupta100% (1)
- Labcir - Marwa - FinalDocument119 pagesLabcir - Marwa - FinalMashavia AhmadNo ratings yet
- EEE111_Multisim_ManualDocument14 pagesEEE111_Multisim_ManualSHADOW manNo ratings yet
- Neil Strauss - The Annihilation Method - Style's Archives - Volume 1 - The Origin of Style - How To Transform From Chump To Champ in No TimeDocument132 pagesNeil Strauss - The Annihilation Method - Style's Archives - Volume 1 - The Origin of Style - How To Transform From Chump To Champ in No TimeDaniel Nii Armah Tetteh100% (2)
- Dear Boy (Acordes para Cancion)Document4 pagesDear Boy (Acordes para Cancion)Rodolfo GuglielmoNo ratings yet
- Number SystemDocument4 pagesNumber SystemGlenn ThomasNo ratings yet
- Max-OT For Dummies - Bodybuilding - Com ForumsDocument21 pagesMax-OT For Dummies - Bodybuilding - Com ForumsPhong QuachNo ratings yet
- SampleContract ShuttleDocument10 pagesSampleContract ShuttleLJ BNo ratings yet
- RD218 Gardan 3110 Ob2226657 NDocument1 pageRD218 Gardan 3110 Ob2226657 Nopen hartigNo ratings yet
- Roof Beam Layout - r1Document1 pageRoof Beam Layout - r1Niraj ShindeNo ratings yet
- Cracking The SQL InterviewDocument52 pagesCracking The SQL InterviewRedouan AFLISSNo ratings yet
- Akuntansi Keuangan Lanjutan - Akuntansi Penggabungan UsahaDocument67 pagesAkuntansi Keuangan Lanjutan - Akuntansi Penggabungan UsahachendyNo ratings yet
- Chemists 12-2023Document7 pagesChemists 12-2023PRC BaguioNo ratings yet
- Safety Data Sheet: Flexa SoftDocument9 pagesSafety Data Sheet: Flexa SoftSercan şahinkayaNo ratings yet
- Complete Tds CourseDocument32 pagesComplete Tds CourseAMLANNo ratings yet
- Fundamentals Deep Learning Activation Functions When To Use ThemDocument15 pagesFundamentals Deep Learning Activation Functions When To Use ThemfaisalNo ratings yet
- WholeDocument397 pagesWholedelbot01No ratings yet
- Irregular Verbs Multiple ChoiceDocument2 pagesIrregular Verbs Multiple Choicecloudcatching0% (1)
- Tugas 3vDocument4 pagesTugas 3vRomie SyafitraNo ratings yet
- Avago Fiber Optic ComponentsDocument8 pagesAvago Fiber Optic ComponentsGerardo BaltaNo ratings yet
- BISE Sukkur HSC Part 2 Results 2015Document12 pagesBISE Sukkur HSC Part 2 Results 2015ajayNo ratings yet
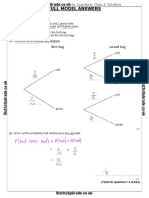
- Probability Tree Diagrams Solutions Mathsupgrade Co UkDocument10 pagesProbability Tree Diagrams Solutions Mathsupgrade Co UknatsNo ratings yet
- Bartending and Catering: Agenda: Basics of Bartending Bar Tools and EquipmentDocument146 pagesBartending and Catering: Agenda: Basics of Bartending Bar Tools and EquipmentMars Mar100% (1)
- Sax AltoDocument2 pagesSax AltoJohnny GervasioNo ratings yet
- 3 Payroll ReportDocument4 pages3 Payroll ReportBen NgNo ratings yet