You might also like
- Tumor MicroenvironmentFrom EverandTumor MicroenvironmentPeter P. LeeNo ratings yet
- OncologistsDocument3 pagesOncologistsAlloush86No ratings yet
- +++kpwilkie Chapter RevisedDocument35 pages+++kpwilkie Chapter RevisedEnes ÇakmakNo ratings yet
- Artículo Citoesqueleto y CáncerDocument17 pagesArtículo Citoesqueleto y CáncerVivian Camila Arguello GonzalezNo ratings yet
- PIIS1074761313004378Document14 pagesPIIS1074761313004378ZadorxsNo ratings yet
- Equipo 12 - Artículo - Fimmu-10-01398Document10 pagesEquipo 12 - Artículo - Fimmu-10-01398Cruz Jardínez Cynthia LuceroNo ratings yet
- On The Discrete Kinetic Theory For Active Particles. Modelling The Immune CompetitionDocument15 pagesOn The Discrete Kinetic Theory For Active Particles. Modelling The Immune CompetitionCarolina RibeiroNo ratings yet
- 8ra19 (2009) Omar A. Ali: Sci Transl Med Et AlDocument12 pages8ra19 (2009) Omar A. Ali: Sci Transl Med Et AlSage RadachowskyNo ratings yet
- The Evolving Tumor Microenvironment From Cancer Initiation To Metastatic OutgrowthDocument30 pagesThe Evolving Tumor Microenvironment From Cancer Initiation To Metastatic OutgrowthJose EdgarNo ratings yet
- Therapeutic Application of T Regulatory Cells in Composite Tissue AllotransplantationDocument15 pagesTherapeutic Application of T Regulatory Cells in Composite Tissue AllotransplantationAngelica SerbulNo ratings yet
- MainDocument16 pagesMainFeby ArdhaniNo ratings yet
- Targeted Therapies to Improve Tumor ImmunotherapyDocument7 pagesTargeted Therapies to Improve Tumor ImmunotherapyMarie ChaisanaNo ratings yet
- Artículo BaseDocument10 pagesArtículo BaseSöö FiiNo ratings yet
- Ru Ella 2013Document25 pagesRu Ella 2013Ruben NatarîșNo ratings yet
- Ascierto1 2013Document4 pagesAscierto1 2013TrabelsiNo ratings yet
- Brain, Behavior, and Immunity Journal Article on Stress and Surgery Promoting Tumor MetastasisDocument10 pagesBrain, Behavior, and Immunity Journal Article on Stress and Surgery Promoting Tumor Metastasisheart wisdomNo ratings yet
- Immunogenic Cell Death in Cancer Review 2019Document12 pagesImmunogenic Cell Death in Cancer Review 2019Kudelko MatNo ratings yet
- Gut Microbiota-MediatedDocument20 pagesGut Microbiota-MediatedMércia FiuzaNo ratings yet
- CIT 2021 Paper 85Document15 pagesCIT 2021 Paper 85Don DiegoNo ratings yet
- Bindea 2014Document3 pagesBindea 2014TrabelsiNo ratings yet
- Jurnal NeoplasmaDocument10 pagesJurnal NeoplasmaFadhilah Asyifa DewantiNo ratings yet
- Celular Dormancy MechanismsDocument17 pagesCelular Dormancy MechanismsJoão FrancoNo ratings yet
- Tumor Immunology: Ineffective in Eliminating Tumors or Preventing Their Growth. (3) Various Means May BeDocument5 pagesTumor Immunology: Ineffective in Eliminating Tumors or Preventing Their Growth. (3) Various Means May BeHillary Desiree RNo ratings yet
- Cancers: Dendritic Cell-Based and Other Vaccination Strategies For Pediatric CancerDocument19 pagesCancers: Dendritic Cell-Based and Other Vaccination Strategies For Pediatric CancerFrendy Ahmad AfandiNo ratings yet
- 3 Tumor ImmunologyDocument12 pages3 Tumor ImmunologyWard KhalidNo ratings yet
- Int Jurnal OncoDocument9 pagesInt Jurnal OncoWida MarianeNo ratings yet
- Harnessing NK Cells To Control MetastasisDocument19 pagesHarnessing NK Cells To Control Metastasisdr. Joko WibowoNo ratings yet
- (24508608 - Journal of Veterinary Research) Mechanisms of Tumour Escape From Immune SurveillanceDocument8 pages(24508608 - Journal of Veterinary Research) Mechanisms of Tumour Escape From Immune Surveillancekalppena naiduNo ratings yet
- Cancerimmunotherapies: Philip J. BergmanDocument22 pagesCancerimmunotherapies: Philip J. BergmanLydia Angelia YanitaNo ratings yet
- Cancerimmunotherapies: Philip J. BergmanDocument22 pagesCancerimmunotherapies: Philip J. BergmanRiefkyansyah PutraNo ratings yet
- Cancer and The Immune System Basic Concepts and Targets For InterventionDocument38 pagesCancer and The Immune System Basic Concepts and Targets For InterventionMariana NannettiNo ratings yet
- Cancer ImmunologyDocument21 pagesCancer ImmunologyYuda PrabawaNo ratings yet
- Review On Cancer and The Immune SystemDocument6 pagesReview On Cancer and The Immune SystemAthenaeum Scientific PublishersNo ratings yet
- Tumor Microenvironment and Immune-Related Therapies of Head and Neck Squamous Cell CarcinomaDocument10 pagesTumor Microenvironment and Immune-Related Therapies of Head and Neck Squamous Cell CarcinomaSpore FluxNo ratings yet
- Monocytes in Tumorigenesis and Tumor ImmunotherapyDocument19 pagesMonocytes in Tumorigenesis and Tumor ImmunotherapyIvan PerezNo ratings yet
- Tumor Immune Escape Mechanisms: Yi Ting Koh,, M. Luz García-Hernández,, and W. Martin KastDocument26 pagesTumor Immune Escape Mechanisms: Yi Ting Koh,, M. Luz García-Hernández,, and W. Martin KastTina HerreraNo ratings yet
- Mauti L JCI 2011Document14 pagesMauti L JCI 2011laetitia.mautiNo ratings yet
- Review Article: Stem Cells: Innovations in Clinical ApplicationsDocument10 pagesReview Article: Stem Cells: Innovations in Clinical Applications03151206639No ratings yet
- The Immune System and Head and Neck Squamous Cell Carcinoma: From Carcinogenesis To New Therapeutic OpportunitiesDocument18 pagesThe Immune System and Head and Neck Squamous Cell Carcinoma: From Carcinogenesis To New Therapeutic OpportunitiesGlauce L TrevisanNo ratings yet
- Reviews: Systemic Immunity in CancerDocument15 pagesReviews: Systemic Immunity in Cancerliliana-contrerasNo ratings yet
- Exosome-Based Immunotherapy: A Promising Approach For Cancer TreatmentDocument16 pagesExosome-Based Immunotherapy: A Promising Approach For Cancer Treatment1810 15Marvin LionelNo ratings yet
- Cancer ModellingDocument15 pagesCancer ModellingAzza Ahmed100% (1)
- Cancer Stem Cell Niche The Place To BeDocument7 pagesCancer Stem Cell Niche The Place To BeAlejandra Isabel Juarez JimenezNo ratings yet
- Mechanisms and Αpplications of Ιnterleukins inDocument21 pagesMechanisms and Αpplications of Ιnterleukins inDefi MarizalNo ratings yet
- Dendritic Cell Gene TherapyDocument16 pagesDendritic Cell Gene TherapyazharbattooNo ratings yet
- Cancer Stem Cell-Immune Cell Crosstalk in Tumor ProgressionDocument11 pagesCancer Stem Cell-Immune Cell Crosstalk in Tumor ProgressionElton De SáNo ratings yet
- Tanyi 2012Document15 pagesTanyi 2012Carlos ImasNo ratings yet
- 1 OnlineDocument14 pages1 OnlinezulalceliikNo ratings yet
- Drake 2006Document3 pagesDrake 2006Ruben NatarîșNo ratings yet
- Inmunoterapia en Cancer de PulmonDocument7 pagesInmunoterapia en Cancer de PulmonJorge ZegarraNo ratings yet
- 2.2. Cancer Immunotherapy Co-Stimulatory Agonists and Co-Inhibitory AntagonistsDocument11 pages2.2. Cancer Immunotherapy Co-Stimulatory Agonists and Co-Inhibitory AntagonistsDiego EskinaziNo ratings yet
- Immunotherapy in Uterine Cancers: Breaking Tolerance and Tilting the Effect Towards Immune SurveillanceDocument9 pagesImmunotherapy in Uterine Cancers: Breaking Tolerance and Tilting the Effect Towards Immune SurveillanceNurul Ulya RahimNo ratings yet
- Mesothelioma Environment Comprises Cytokines and T-Regulatory Cells That Suppress Immune ResponsesDocument10 pagesMesothelioma Environment Comprises Cytokines and T-Regulatory Cells That Suppress Immune ResponseshariompriyNo ratings yet
- Managing The Immune Microenvironment of Osteosarcoma - The Outlook For Osteosarcoma TreatmentDocument26 pagesManaging The Immune Microenvironment of Osteosarcoma - The Outlook For Osteosarcoma TreatmentoudyNo ratings yet
- Antibody-Cytokine Fusion ProteinsDocument9 pagesAntibody-Cytokine Fusion ProteinsShima YousefiNo ratings yet
- Current Status and Future Directions of Cancer ImmunotherapyDocument9 pagesCurrent Status and Future Directions of Cancer ImmunotherapyRo KohnNo ratings yet
- Host Defence Mechanisms Against TumorsDocument7 pagesHost Defence Mechanisms Against TumorsHamam AmastrdamNo ratings yet
- Paper2005 PDFDocument4 pagesPaper2005 PDFJoshua GatesNo ratings yet
- Immune Response in The Skin of AgingDocument7 pagesImmune Response in The Skin of AgingInmunoBlogNo ratings yet
- Challenges in The Development of Immunoisolation DevicesDocument16 pagesChallenges in The Development of Immunoisolation DevicesGarret WilliamsNo ratings yet
- Astm A 322 - 07Document4 pagesAstm A 322 - 07Luis MendozaNo ratings yet
- Mic10 PDFDocument2 pagesMic10 PDFArnold c ElverNo ratings yet
- Ecaadesigradi2019 465Document10 pagesEcaadesigradi2019 465Rhino ArdiawanNo ratings yet
- MSM Module 1Document59 pagesMSM Module 1Nk KumarNo ratings yet
- 1.3 Errors and UncertaintyDocument24 pages1.3 Errors and UncertaintyHerman HermanNo ratings yet
- Year 9 Core Autumn B 2021Document16 pagesYear 9 Core Autumn B 2021sheikhaalfahdiNo ratings yet
- The Dynamic Characteristics Analysis of Rotor Blade Based On ANSYSDocument11 pagesThe Dynamic Characteristics Analysis of Rotor Blade Based On ANSYSFrancisco HidalgoNo ratings yet
- Cambridge O Level: Mathematics (Syllabus D) 4024/22Document20 pagesCambridge O Level: Mathematics (Syllabus D) 4024/22D.T.CNo ratings yet
- Ada Villafafila Garcia, Ph.d.Document249 pagesAda Villafafila Garcia, Ph.d.Alberto Vázquez ContrerasNo ratings yet
- Sample Computation For FootingDocument3 pagesSample Computation For FootingDenice CastroNo ratings yet
- Photodiode & Solar Cell FundamentalsDocument33 pagesPhotodiode & Solar Cell Fundamentalsgirishkumardarisi254No ratings yet
- Thermal Bridge DisplacementsDocument18 pagesThermal Bridge DisplacementsAhmed KhalafNo ratings yet
- Design of T BeamDocument5 pagesDesign of T BeamAnton_Young_1962No ratings yet
- Mud Logger # 10Document8 pagesMud Logger # 10Aiham AltayehNo ratings yet
- Profil Paparan Radiasi Instalasi Radiologi - Dwi RochmayantiDocument5 pagesProfil Paparan Radiasi Instalasi Radiologi - Dwi RochmayantiFadila Nur'AeniNo ratings yet
- Pascua Project For Module 2 Problem Set With SolutionDocument14 pagesPascua Project For Module 2 Problem Set With SolutionDJ KillNo ratings yet
- Activity 3: Bjective Aterial EquiredDocument3 pagesActivity 3: Bjective Aterial EquiredMalati MauryaNo ratings yet
- Electromagnetic Spectrum and Optics AssessmentDocument5 pagesElectromagnetic Spectrum and Optics AssessmentKenneth Roy MatuguinaNo ratings yet
- Guide to Solving Differential EquationsDocument8 pagesGuide to Solving Differential Equationsسعد احمد حميديNo ratings yet
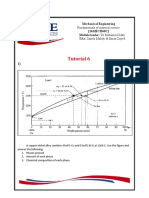
- Tutorial 6: Mechanical Engineering (16Mech60C) Module Leader: DR - Mohamed Zaky Tas: Sameh Mahdy & Eman ZayedDocument3 pagesTutorial 6: Mechanical Engineering (16Mech60C) Module Leader: DR - Mohamed Zaky Tas: Sameh Mahdy & Eman Zayedsherif115040 BueNo ratings yet
- SJI Technical Digest 11 Jan2021 Pf871rDocument98 pagesSJI Technical Digest 11 Jan2021 Pf871rCAROL REYES LLUBERESNo ratings yet
- EBM Axial - EN - Mini - 070921Document108 pagesEBM Axial - EN - Mini - 070921a ferreira e oliveiraNo ratings yet
- Introduction to Solid Modeling Using SolidWorks 2012 SolidWorks Simulation TutorialDocument8 pagesIntroduction to Solid Modeling Using SolidWorks 2012 SolidWorks Simulation TutorialAndi SuntoroNo ratings yet
- Light Emitting Diodes: Figure 1: Schematic Diagram of An LEDDocument9 pagesLight Emitting Diodes: Figure 1: Schematic Diagram of An LEDVikash SingleNo ratings yet
- Saint Ferdinand College Sta. Ana Street, City of Ilagan, Isabela Senior High School DepartmentDocument5 pagesSaint Ferdinand College Sta. Ana Street, City of Ilagan, Isabela Senior High School DepartmentAngela AlejandroNo ratings yet
- Upthrust and Archimedes Principle Moments and Equilibrium PDFDocument11 pagesUpthrust and Archimedes Principle Moments and Equilibrium PDFAlisa DowningNo ratings yet
- Hypothesis Development: Module I - Topic VDocument12 pagesHypothesis Development: Module I - Topic VLissetteMachadoNo ratings yet
- ATOMIC STRUCTURE NOTES FOR CLASS 11Document15 pagesATOMIC STRUCTURE NOTES FOR CLASS 11Bhavya Goyal XI Non med91% (11)
- GIET UNIVERSITY PARTIAL DIFFERENTIAL EQUATIONS EXAMDocument2 pagesGIET UNIVERSITY PARTIAL DIFFERENTIAL EQUATIONS EXAMPritesh RayaguruNo ratings yet
- The Ins and Outs of Human Cognition in The Construction of MeaningDocument11 pagesThe Ins and Outs of Human Cognition in The Construction of MeaningmarceloNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)