You might also like
- Unit2 Molecular Bio QuestionsDocument128 pagesUnit2 Molecular Bio QuestionsSıla DenizNo ratings yet
- Non-Glycolytic Pathways of Metabolism of GlucoseFrom EverandNon-Glycolytic Pathways of Metabolism of GlucoseRating: 1 out of 5 stars1/5 (1)
- Metabolism of Lipids: The Biochemistry and Molecular Biology Department of CMUDocument270 pagesMetabolism of Lipids: The Biochemistry and Molecular Biology Department of CMUanon_49132843No ratings yet
- Chapter 18 Metabolic PathwaysDocument133 pagesChapter 18 Metabolic PathwaysM3hdi87No ratings yet
- Mitochondria Cellular RespirationDocument65 pagesMitochondria Cellular Respirationpotski413No ratings yet
- GluconeogenesisDocument31 pagesGluconeogenesisRajakannanNo ratings yet
- Homeostasis: (Link)Document3 pagesHomeostasis: (Link)Shereen AlobinayNo ratings yet
- Physical Organic Chemistry—Ii: Specially Invited Lectures Presented at the Second IUPAC Conference on Physical Organic Chemistry Held at Noordwijkerhout, Netherlands, 29 April–2 May 1974From EverandPhysical Organic Chemistry—Ii: Specially Invited Lectures Presented at the Second IUPAC Conference on Physical Organic Chemistry Held at Noordwijkerhout, Netherlands, 29 April–2 May 1974Th. J. De BoerNo ratings yet
- Chapter 27 BioenergeticsDocument44 pagesChapter 27 BioenergeticsELAINE MARIEJOY MANGAOILNo ratings yet
- Biochem 1.5 Bioenergetics PDFDocument7 pagesBiochem 1.5 Bioenergetics PDFlovelots1234No ratings yet
- Primary and Secondary MetabolitesDocument27 pagesPrimary and Secondary Metabolitesshubham sabaleNo ratings yet
- EFE & CPM of FordDocument4 pagesEFE & CPM of FordShereen AlobinayNo ratings yet
- Bioenergetics How The Body Converts Food To EnergyDocument38 pagesBioenergetics How The Body Converts Food To EnergyShereen AlobinayNo ratings yet
- Nursing TheoristsDocument92 pagesNursing TheoristsShereen AlobinayNo ratings yet
- Fatty Acid Synthesis 11.12.19Document18 pagesFatty Acid Synthesis 11.12.19Sanreet Randhawa100% (1)
- FullDocument1,497 pagesFullSyasa PembayunNo ratings yet
- CHEM F343: Inorganic Chemistry III: PilaniDocument29 pagesCHEM F343: Inorganic Chemistry III: PilaniANANYA PUPNEJANo ratings yet
- Cells and Sugars 7-Gluconeogenesis-StudentDocument15 pagesCells and Sugars 7-Gluconeogenesis-StudenttyhbbhhNo ratings yet
- Cellular Respiration: Glycolytic PathwaysDocument34 pagesCellular Respiration: Glycolytic PathwaysInshirah IzhamNo ratings yet
- Degradation of TG: 1 - Fat catabolism (lipolysis) 2.β-Oxidation of Fatty acids (fatty acid oxidation)Document26 pagesDegradation of TG: 1 - Fat catabolism (lipolysis) 2.β-Oxidation of Fatty acids (fatty acid oxidation)Aboubakar Moalim Mahad moh'dNo ratings yet
- Carbohydrate Metabolism 1Document70 pagesCarbohydrate Metabolism 1nia adelleNo ratings yet
- Chapter 14 BiomoleculesDocument11 pagesChapter 14 BiomoleculesJaanvi SisodiaNo ratings yet
- Carbohydrate Metabolism 02-The FATE of PYRUVATEDocument22 pagesCarbohydrate Metabolism 02-The FATE of PYRUVATEokolodivine334No ratings yet
- Fatty Acid Oxidation and Synthesis of Ketone BodyDocument58 pagesFatty Acid Oxidation and Synthesis of Ketone BodyCeciliaNo ratings yet
- Struktur, Fungsi Dan Metabolisme Karbohidrat: Department of Biochemistryy Medical Faculti of Hasanuddin UniversityDocument103 pagesStruktur, Fungsi Dan Metabolisme Karbohidrat: Department of Biochemistryy Medical Faculti of Hasanuddin UniversityNicha gunawanfarizalNo ratings yet
- 5 Carbohydrate Metabolism LectureDocument53 pages5 Carbohydrate Metabolism LectureAsrorNo ratings yet
- Glycolysis, Pyruvate Catabolism, Citric Acid CycleDocument8 pagesGlycolysis, Pyruvate Catabolism, Citric Acid CycleIssacNo ratings yet
- Lipida II2008Document54 pagesLipida II2008Dika Virga SaputraNo ratings yet
- Biology Assignment - 20231106 - 003633 - 0000Document17 pagesBiology Assignment - 20231106 - 003633 - 0000vikashjejusar1No ratings yet
- ProstaglandinsDocument43 pagesProstaglandinsRandy BrownNo ratings yet
- Specific Catabolic PathwaysDocument28 pagesSpecific Catabolic PathwaysSiddarth PalletiNo ratings yet
- Glycolysis GluconeogenesisDocument66 pagesGlycolysis GluconeogenesisRahmad AllulNo ratings yet
- AA BiosynthesisDocument69 pagesAA BiosynthesisSajid NabiNo ratings yet
- Lecture 10 - GlycolysisDocument30 pagesLecture 10 - GlycolysisSyafiqah SuhaimiNo ratings yet
- Biosynthetic Pathways: Chemistry 203Document50 pagesBiosynthetic Pathways: Chemistry 203Jake OteroNo ratings yet
- Lipid MetabolismDocument81 pagesLipid MetabolismnurwahidahNo ratings yet
- + F.R.Fatty Acid SynthesisDocument28 pages+ F.R.Fatty Acid SynthesisAmanuel MaruNo ratings yet
- 12 - Chemistry - Notes - ch14 - Biomolecules (2) Word FormDocument12 pages12 - Chemistry - Notes - ch14 - Biomolecules (2) Word FormSurekha raoNo ratings yet
- TransaminationDocument19 pagesTransaminationMehak shaikh50% (2)
- Photorespiratio NDocument11 pagesPhotorespiratio NSheelendra Mangal BhattNo ratings yet
- EnergyDocument34 pagesEnergymoral alabsioNo ratings yet
- Lipid & AA MetabolismDocument90 pagesLipid & AA MetabolismUmi LatifahNo ratings yet
- Photosynthesis: Prof. S. Dutta GuptaDocument41 pagesPhotosynthesis: Prof. S. Dutta Guptahimanshu_agraNo ratings yet
- Fatty Acid OxidationDocument43 pagesFatty Acid OxidationAmalia DarwisNo ratings yet
- Carbohydrate MetabolismDocument72 pagesCarbohydrate MetabolismJeffson BalmoresNo ratings yet
- Specific Catabolic Pathways: Carbohydrate, Lipid, and Protein MetabolismDocument43 pagesSpecific Catabolic Pathways: Carbohydrate, Lipid, and Protein MetabolismShereen AlobinayNo ratings yet
- GOB9e - Chapter28 - Metabolism Carbo, Lipids, ProteinsDocument43 pagesGOB9e - Chapter28 - Metabolism Carbo, Lipids, Proteinsasieee chimmyNo ratings yet
- Respiration-Glycolysis and Krebs CycleDocument17 pagesRespiration-Glycolysis and Krebs CycleTwinkle MounamiNo ratings yet
- Catabolic Pathways Chapter28Document45 pagesCatabolic Pathways Chapter28Reena Mae GomezNo ratings yet
- Chapter Viii - Carbohydrates MechanismDocument34 pagesChapter Viii - Carbohydrates MechanismAngelo AngelesNo ratings yet
- Fatty Acid Degradation and Biosynthesis: Department of BiochemistryDocument55 pagesFatty Acid Degradation and Biosynthesis: Department of Biochemistrydentcu70No ratings yet
- 1) Absorption of Carbohydrates:: The Small Intestine Is The Main Absorptive Organ (90% of Ingested Foodstuffs & H O)Document13 pages1) Absorption of Carbohydrates:: The Small Intestine Is The Main Absorptive Organ (90% of Ingested Foodstuffs & H O)api-3706483No ratings yet
- Oxidation of Fatty AcidsDocument90 pagesOxidation of Fatty AcidsDeepa DevanathanNo ratings yet
- PhotosynthesisDocument55 pagesPhotosynthesischezkanoelleangeliebalolongNo ratings yet
- 22 CH106 Metabolic Paths For Carbohydrates Timberlake 2ndDocument70 pages22 CH106 Metabolic Paths For Carbohydrates Timberlake 2ndEnrique LiKeNo ratings yet
- Chapter 6 - Photosynthesis Carbon MetabolismDocument46 pagesChapter 6 - Photosynthesis Carbon Metabolismsayidah nafisahNo ratings yet
- Dark Reaction Topics: CO Fixation Calvin Cycle Starch and Sucrose BiosynthesisDocument14 pagesDark Reaction Topics: CO Fixation Calvin Cycle Starch and Sucrose BiosynthesisHamza SarrajNo ratings yet
- Metabolic Pathways For CarbohydratesDocument15 pagesMetabolic Pathways For CarbohydratesLa Ode RinaldiNo ratings yet
- 07 - Glycolysis, Tca Cycle and Oxidative PhosphorylationDocument52 pages07 - Glycolysis, Tca Cycle and Oxidative PhosphorylationhabibahNo ratings yet
- Biochemistry of The EyeDocument38 pagesBiochemistry of The EyeYusuf Almalik SaputraNo ratings yet
- Glycolysis and Gluconeogenesis: Instructor-RclDocument33 pagesGlycolysis and Gluconeogenesis: Instructor-RclNitish ShahNo ratings yet
- Carbohydrate Metabolism + Analysis of CarbohydratesDocument51 pagesCarbohydrate Metabolism + Analysis of CarbohydratesRara SorayaNo ratings yet
- Digestion of Triacylglycerols Beta-Oxidation of Fatty Acids ATP and Fatty Acid OxidationDocument31 pagesDigestion of Triacylglycerols Beta-Oxidation of Fatty Acids ATP and Fatty Acid OxidationNICHAEL MARIA CELINA UYNo ratings yet
- LECT 22 FAsynthesisDocument21 pagesLECT 22 FAsynthesisjoko16No ratings yet
- Metabolism and Energy ProductionDocument66 pagesMetabolism and Energy ProductionDrick AsiloNo ratings yet
- Intro To Metabolism Glycolysis Krebs Cycle ETCDocument104 pagesIntro To Metabolism Glycolysis Krebs Cycle ETCAnna Sofia ReyesNo ratings yet
- Gluco Neo GenesisDocument36 pagesGluco Neo GenesisV sNo ratings yet
- Food Metabolism: Chemistry DepartmentDocument33 pagesFood Metabolism: Chemistry DepartmentMirawati EfendiNo ratings yet
- FACTORS AFFECTING GROWTH AND DEVELOPMENT by NavarroDocument5 pagesFACTORS AFFECTING GROWTH AND DEVELOPMENT by NavarroShereen AlobinayNo ratings yet
- Basics of Human Genetics - AMBOSSDocument21 pagesBasics of Human Genetics - AMBOSSShereen AlobinayNo ratings yet
- Specific Catabolic Pathways: Carbohydrate, Lipid, and Protein MetabolismDocument43 pagesSpecific Catabolic Pathways: Carbohydrate, Lipid, and Protein MetabolismShereen AlobinayNo ratings yet
- Immunochemistry: Frederick A. Bettelheim William H. Brown Mary K. Campbell Shawn O. FarrellDocument41 pagesImmunochemistry: Frederick A. Bettelheim William H. Brown Mary K. Campbell Shawn O. FarrellShereen AlobinayNo ratings yet
- Nutrition: Frederick A. Bettelheim William H. Brown Mary K. Campbell Shawn O. FarrellDocument15 pagesNutrition: Frederick A. Bettelheim William H. Brown Mary K. Campbell Shawn O. FarrellShereen AlobinayNo ratings yet
- Anatomy - Endocrine System - HormonesDocument21 pagesAnatomy - Endocrine System - HormonesShereen AlobinayNo ratings yet
- IE MatrixDocument5 pagesIE MatrixShereen AlobinayNo ratings yet
- The Price Tag of Leadership: Self-DisciplineDocument14 pagesThe Price Tag of Leadership: Self-DisciplineShereen AlobinayNo ratings yet
- Intake and OutputDocument3 pagesIntake and OutputShereen AlobinayNo ratings yet
- Mitigation and Responsiveness of Philippines Government Towards Natural CalamitiesDocument1 pageMitigation and Responsiveness of Philippines Government Towards Natural CalamitiesShereen AlobinayNo ratings yet
- Adolescent and Youth Health Program (AYHP)Document11 pagesAdolescent and Youth Health Program (AYHP)Shereen AlobinayNo ratings yet
- Filipino GamesDocument5 pagesFilipino GamesShereen AlobinayNo ratings yet
- General Urine Specimen CollectionDocument3 pagesGeneral Urine Specimen CollectionShereen AlobinayNo ratings yet
- Philippine Maguindanao Maranao Armampents: Sagayan Festival (Maranao War Dance)Document2 pagesPhilippine Maguindanao Maranao Armampents: Sagayan Festival (Maranao War Dance)Shereen AlobinayNo ratings yet
- Filipino Nationalism AcceleratorsDocument9 pagesFilipino Nationalism AcceleratorsShereen AlobinayNo ratings yet
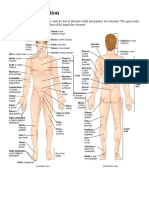
- Anatomy - Skeletal SystemDocument26 pagesAnatomy - Skeletal SystemShereen AlobinayNo ratings yet
- Tripalmitin Names: Formula Molecular WeightDocument4 pagesTripalmitin Names: Formula Molecular WeightShereen AlobinayNo ratings yet
- Bedpan or A Urinal: What You Need To Know: What Are Bedpans and Urinals?Document8 pagesBedpan or A Urinal: What You Need To Know: What Are Bedpans and Urinals?Shereen AlobinayNo ratings yet
- Psychology - OVERVIEWDocument13 pagesPsychology - OVERVIEWShereen AlobinayNo ratings yet
- Anatomical TerminologyDocument5 pagesAnatomical TerminologyShereen AlobinayNo ratings yet
- Introductory Biochemistry: CHEM 160Document31 pagesIntroductory Biochemistry: CHEM 160Gianna Kristen MirandaNo ratings yet
- Gaudy - Microbiology For Environmental Scientists and EngineersDocument753 pagesGaudy - Microbiology For Environmental Scientists and EngineersEnes GiderNo ratings yet
- NCERT Solution For Class 11 Biology Chapter 14 Respiration in PlantsDocument6 pagesNCERT Solution For Class 11 Biology Chapter 14 Respiration in PlantsHafiz HussainNo ratings yet
- METABOLISM of CARBOHYDRATESDocument62 pagesMETABOLISM of CARBOHYDRATESmohaysin pirinoNo ratings yet
- Metabolism: Anabolism and CatabolismDocument4 pagesMetabolism: Anabolism and CatabolismMedi OmicNo ratings yet
- Module 3 - Finals - Introduction To Microbial Metabolism-1Document40 pagesModule 3 - Finals - Introduction To Microbial Metabolism-1Mary Grace ClarosNo ratings yet
- Biomolecules Crash Course NotesDocument22 pagesBiomolecules Crash Course NotesAayush sainiNo ratings yet
- Carbohydrate Metabolism in PlantsDocument40 pagesCarbohydrate Metabolism in Plantsguswinda alvaniNo ratings yet
- What To Never Eat After Your Work OutDocument39 pagesWhat To Never Eat After Your Work OutDimitar FilevskiNo ratings yet
- Unit I I Carbohydrates MetabolismDocument33 pagesUnit I I Carbohydrates Metabolismseno adi100% (2)
- This Is A SampleDocument10 pagesThis Is A SampleNashi TagazaNo ratings yet
- From Metabolomics To FluxomicsDocument10 pagesFrom Metabolomics To FluxomicscamartinezriNo ratings yet
- As BiomethylationDocument5 pagesAs BiomethylationZAIB KOHLINo ratings yet
- Download pdf Essential Biochemistry Kathleen Cornely ebook full chapterDocument53 pagesDownload pdf Essential Biochemistry Kathleen Cornely ebook full chapterjames.kirby133100% (1)
- Bms2 Integration of MetabolismDocument42 pagesBms2 Integration of MetabolismhohoylandNo ratings yet
- Biodegradacion de XenobioticosDocument8 pagesBiodegradacion de XenobioticosDoc Efra RamirezNo ratings yet
- NMAT Lec 2 Organic ChemistryDocument121 pagesNMAT Lec 2 Organic ChemistryMikaela Rome BigayNo ratings yet
- Adenosine Triphosphate - WikipediaDocument65 pagesAdenosine Triphosphate - WikipediaBashiir NuurNo ratings yet
- FATTY ACID OXIDATION (Beta-Oxidation) ...Document9 pagesFATTY ACID OXIDATION (Beta-Oxidation) ...atifzeaNo ratings yet
- Erythrocyte Metabolism AND Membrane Structure and FunctionDocument29 pagesErythrocyte Metabolism AND Membrane Structure and FunctionJohn Marie IdavaNo ratings yet
- Gpat Grand TestDocument12 pagesGpat Grand TestGANESH KUMAR JELLA100% (2)
- Acetate PathwayDocument25 pagesAcetate PathwaySangga Buana KomaraNo ratings yet
- Nutrition & Bio-Chemistry PDFDocument10 pagesNutrition & Bio-Chemistry PDFRãj Pôtti ÑâîkNo ratings yet
- Phospho Ketolase PathwayDocument2 pagesPhospho Ketolase PathwayDr. SHIVA AITHALNo ratings yet
- Integration of MetabolismDocument10 pagesIntegration of MetabolismAditya NayakNo ratings yet
- BMSC 230 Online Quizzes Module 4-6Document12 pagesBMSC 230 Online Quizzes Module 4-6Brennan J.G. Kumitch0% (1)