You might also like
- HHS Public Access: Lyme DiseaseDocument14 pagesHHS Public Access: Lyme DiseasemikiNo ratings yet
- Lyme Disease SpeechDocument4 pagesLyme Disease SpeechDominic SteigmajerNo ratings yet
- NIH Public Access: Lyme DiseaseDocument25 pagesNIH Public Access: Lyme DiseasemikiNo ratings yet
- Lyme DiseaseDocument4 pagesLyme DiseaseAyub MaingiNo ratings yet
- Lyme Disease in Dogs and Cats Guide: Symptoms, Tests and TreatmentsDocument17 pagesLyme Disease in Dogs and Cats Guide: Symptoms, Tests and TreatmentskerokunNo ratings yet
- Japanese Encephalitis: Neurological Aspects of Tropical DiseaseDocument11 pagesJapanese Encephalitis: Neurological Aspects of Tropical DiseaseserubimNo ratings yet
- Japanese Encephalitis: Neurological Aspects of Tropical DiseaseDocument11 pagesJapanese Encephalitis: Neurological Aspects of Tropical Diseaseyasaswini royNo ratings yet
- Arboviruses: Robert E. Shope and James M. MeeganDocument2 pagesArboviruses: Robert E. Shope and James M. Meeganfelipe andreNo ratings yet
- 32 Arthropod Borne DiseasesDocument16 pages32 Arthropod Borne Diseasesvieira.barrosNo ratings yet
- Yellow FeverDocument9 pagesYellow Feversag50942723No ratings yet
- Landmark 2146Document13 pagesLandmark 2146y2h996tnp8No ratings yet
- Australia's War Against Rabbits: The Story of Rabbit Haemorrhagic DiseaseFrom EverandAustralia's War Against Rabbits: The Story of Rabbit Haemorrhagic DiseaseNo ratings yet
- Epidemic Respiratory Disease: Pneumonias & Other Infections Accompanying Influenza and MeaslesFrom EverandEpidemic Respiratory Disease: Pneumonias & Other Infections Accompanying Influenza and MeaslesNo ratings yet
- Solomon 2000Document12 pagesSolomon 2000nathanaellee92No ratings yet
- Summary of Suzanne Humphries & Roman Bystrianyk's Dissolving IllusionsFrom EverandSummary of Suzanne Humphries & Roman Bystrianyk's Dissolving IllusionsNo ratings yet
- Ebola Virus Disease: A Manual for EVD ManagementFrom EverandEbola Virus Disease: A Manual for EVD ManagementMarta LadoNo ratings yet
- 1986 MS Caused by Lyme US StudyDocument9 pages1986 MS Caused by Lyme US StudyTegorion WhiteNo ratings yet
- Lyme DiseaseDocument12 pagesLyme Diseaseflo1234roNo ratings yet
- Epidemic, Pandemic, Should I Call the Medic? Biology Books for Kids | Children's Biology BooksFrom EverandEpidemic, Pandemic, Should I Call the Medic? Biology Books for Kids | Children's Biology BooksNo ratings yet
- Blue Tongue by NabeelDocument115 pagesBlue Tongue by NabeelDr Anais AsimNo ratings yet
- Babesia FinalDocument104 pagesBabesia FinalSameerPeerNo ratings yet
- Elewski 2000Document20 pagesElewski 2000mgalihwbNo ratings yet
- Scrub TyphusDocument6 pagesScrub Typhusjinsi georgeNo ratings yet
- Curs 14 Boala Lyme 2Document63 pagesCurs 14 Boala Lyme 2n1u1s1h1100% (1)
- Who Is This: FlaviviridaeDocument10 pagesWho Is This: FlaviviridaeCarlton RanjithNo ratings yet
- Lancet Review Yellow Fever 2001Document10 pagesLancet Review Yellow Fever 20011997finalfantasyNo ratings yet
- Leptospirosis Cap 142Document11 pagesLeptospirosis Cap 142EdsonPerezNo ratings yet
- Emerging and ReDocument5 pagesEmerging and RedewikurnialestariNo ratings yet
- Little2010 (Lyme Borreliosis in Dogs and Humans in The USA)Document6 pagesLittle2010 (Lyme Borreliosis in Dogs and Humans in The USA)KC ONo ratings yet
- Enfermedad de Lyme (Neuroborreliosis) - ContinuumDocument16 pagesEnfermedad de Lyme (Neuroborreliosis) - ContinuumNeurología RebagliatiNo ratings yet
- Bronquite Infecciosa EuropaDocument4 pagesBronquite Infecciosa EuropaRafaellaCalzadaNo ratings yet
- Rabies in Small Animals: Wildlife Threat and Clinical SignsDocument11 pagesRabies in Small Animals: Wildlife Threat and Clinical Signssindika anastasyaNo ratings yet
- BICOM Treatment of Chronic Fatigue and FibromyalgiaCFS ReportDocument15 pagesBICOM Treatment of Chronic Fatigue and FibromyalgiaCFS ReportAnahata Fraydon100% (1)
- Brenda Alvarez Leccion 1Document5 pagesBrenda Alvarez Leccion 1Brenda AlvNo ratings yet
- HHS Public Access: 5. Diagnosis and Treatment of Lyme ArthritisDocument15 pagesHHS Public Access: 5. Diagnosis and Treatment of Lyme ArthritismikiNo ratings yet
- Update on yellow fever virus: epidemiology, transmission, clinical features and preventionDocument10 pagesUpdate on yellow fever virus: epidemiology, transmission, clinical features and preventionRoy MontoyaNo ratings yet
- Lyme Disease Epidemic Caused by Human Disruption of EnvironmentDocument8 pagesLyme Disease Epidemic Caused by Human Disruption of EnvironmentHelena TakahashiNo ratings yet
- 408 2022 Article 528Document8 pages408 2022 Article 528nathanaellee92No ratings yet
- Lyme Borreliosis in Europe and North America: Epidemiology and Clinical PracticeFrom EverandLyme Borreliosis in Europe and North America: Epidemiology and Clinical PracticeSunil K. SoodNo ratings yet
- 7 Deadliest Diseases in HistoryDocument9 pages7 Deadliest Diseases in HistoryRth CMglNo ratings yet
- Using The Flea As A WeaponDocument6 pagesUsing The Flea As A WeaponReid KirbyNo ratings yet
- Cowpox Jenner 1996Document2 pagesCowpox Jenner 1996Chrislyn SanlaoNo ratings yet
- Covid: Viruses/Vaccines—: Covid-19 Outbreak Worldwide PandemicFrom EverandCovid: Viruses/Vaccines—: Covid-19 Outbreak Worldwide PandemicNo ratings yet
- Cambridge AS Level Biology Section: I Infectious DiseasesDocument12 pagesCambridge AS Level Biology Section: I Infectious Diseaseswaheed aluNo ratings yet
- Region III HIV/AIDS Cases Rise in 2017Document8 pagesRegion III HIV/AIDS Cases Rise in 2017MHO Ng DRT BulacanNo ratings yet
- Free Sample Informative SpeechDocument1 pageFree Sample Informative SpeechSari Adnin HalimNo ratings yet
- Sexually Transmitted Diseases: Case Studies 2015 WV Public Health SymposiumDocument28 pagesSexually Transmitted Diseases: Case Studies 2015 WV Public Health SymposiumNikole CrastaNo ratings yet
- Ebola Virus Oet Reading Subtest PracticeDocument4 pagesEbola Virus Oet Reading Subtest PracticeSanjana SabuNo ratings yet
- EbolaDocument15 pagesEbolaIbama MirillaNo ratings yet
- Common RashesDocument73 pagesCommon Rashesmay0s910No ratings yet
- Vector Borne Diseases: Presentator: Atikah Najla RodhiahDocument15 pagesVector Borne Diseases: Presentator: Atikah Najla RodhiahTengku Mossadeq Al QornyNo ratings yet
- Hand Foot and Mouth DiseaseDocument16 pagesHand Foot and Mouth DiseaseCaroline TecsonNo ratings yet
- S2-Epid - Main Disese (3) - Influenza - Student - 2023Document48 pagesS2-Epid - Main Disese (3) - Influenza - Student - 2023Syahrul SanmasNo ratings yet
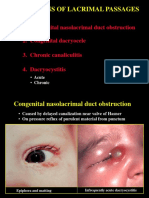
- Infections of Lacrimal PassagesDocument8 pagesInfections of Lacrimal PassagesgeorginaNo ratings yet
- Rubella and PregnancyDocument6 pagesRubella and PregnancyKABERA RENENo ratings yet
- African Swine Fever: A Highly Contagious Hemorrhagic DiseaseDocument11 pagesAfrican Swine Fever: A Highly Contagious Hemorrhagic DiseaseTamiru LegesseNo ratings yet
- CoronavirusDocument3 pagesCoronavirusĂhméđ ĂĺãshqãrNo ratings yet
- Malaria Leaflet PDFDocument2 pagesMalaria Leaflet PDFPrimus moneNo ratings yet
- Gilut Herpes ZosteRDocument24 pagesGilut Herpes ZosteRdimasahadiantoNo ratings yet
- Jurnal KKN - Singgih Dwi Prasetyo UNSDocument11 pagesJurnal KKN - Singgih Dwi Prasetyo UNSSinggih Dwi PrasetyoNo ratings yet
- What is hepatitis B virus infection and how is it treatedDocument2 pagesWhat is hepatitis B virus infection and how is it treatedVictor Iv MirandaNo ratings yet
- Bird FluDocument5 pagesBird FluNader SmadiNo ratings yet
- Icd 10 20052Document380 pagesIcd 10 20052Duff DuffNo ratings yet
- NCM 104 Lec Syllabus MELDocument5 pagesNCM 104 Lec Syllabus MELLanz LlnsNo ratings yet
- Notable Pandemics in HistoryDocument2 pagesNotable Pandemics in Historyapi-264004571No ratings yet
- ON Plasmodium Vivax (Malarial Parasite)Document10 pagesON Plasmodium Vivax (Malarial Parasite)Raja Marudut SimatupangNo ratings yet
- Cleft Lip and PalateDocument50 pagesCleft Lip and Palatesusanti bulanNo ratings yet
- Dapus FixDocument2 pagesDapus FixIkrima MuhdarmuhallyNo ratings yet
- Daftar Pustaka: of Life. 44. Journal of Pediatric Surgery, 2009Document3 pagesDaftar Pustaka: of Life. 44. Journal of Pediatric Surgery, 2009Lela Nazila ItuSayaNo ratings yet
- Tinea Cruris Abq JournalDocument3 pagesTinea Cruris Abq JournalGustiandari FidhyaNo ratings yet
- Introduction To MycologyDocument66 pagesIntroduction To MycologyHerlinaNababanNo ratings yet
- Literature Review of Swine FluDocument5 pagesLiterature Review of Swine Fluc5qd4nn8100% (1)
- Gallimune 407 ND+IB+EDS+ART: PresentationDocument2 pagesGallimune 407 ND+IB+EDS+ART: PresentationRahmat SyahrirNo ratings yet