You might also like
- Smokable DMT from Organic SourcesDocument12 pagesSmokable DMT from Organic SourcesRolando Ross100% (1)
- British Journal of Nutrition (2008), 99, E-Suppl.Document50 pagesBritish Journal of Nutrition (2008), 99, E-Suppl.nihadtoussounNo ratings yet
- Treatment and Management of Mental Health Conditions During Pregnancy and Postpartum ACOG June 2023Document27 pagesTreatment and Management of Mental Health Conditions During Pregnancy and Postpartum ACOG June 2023kaiorm2k100% (3)
- Medicinal Chemistry (MCQs Exam)Document10 pagesMedicinal Chemistry (MCQs Exam)AbCc100% (7)
- Target Identification and ValidationDocument21 pagesTarget Identification and ValidationManuel Christopher MontesclarosNo ratings yet
- Antioxidant 2Document8 pagesAntioxidant 2Gayathri deviNo ratings yet
- Ecam2015 620472Document14 pagesEcam2015 620472FrengkyNo ratings yet
- TubaDocument11 pagesTubaAnnisa Dian0% (1)
- Antimicrobial Activity of FlavonoidsDocument27 pagesAntimicrobial Activity of Flavonoidsmuntiir gurusingaNo ratings yet
- Patel-Goyal2012 Article RecentDevelopmentsInMushroomsADocument15 pagesPatel-Goyal2012 Article RecentDevelopmentsInMushroomsAalNo ratings yet
- NPR Usnic Acid Review 2015Document17 pagesNPR Usnic Acid Review 2015Restuan Lubis SudirmanNo ratings yet
- Acacia Mangium Willd (Acácia) Extract Antioxidant and Antiproliferative ActivitiesDocument10 pagesAcacia Mangium Willd (Acácia) Extract Antioxidant and Antiproliferative ActivitiesSabrina JonesNo ratings yet
- MMMDocument10 pagesMMMWiri Resky AmaliaNo ratings yet
- Methods For Total Antioxidant Activity Determination A Review 2161 1009.1000106Document10 pagesMethods For Total Antioxidant Activity Determination A Review 2161 1009.1000106Sie ningsihNo ratings yet
- Liu - 2011 - The Antioxidant and Free-Radical Scavenging Activities of Extract and Fractions PDFDocument9 pagesLiu - 2011 - The Antioxidant and Free-Radical Scavenging Activities of Extract and Fractions PDFAndika Yuli HeryantoNo ratings yet
- Secondary Metabolism and Antimicrobial Metabolites of AspergillusDocument13 pagesSecondary Metabolism and Antimicrobial Metabolites of AspergillusBrianNo ratings yet
- JPAM Vol 15 Issue1 P 285-299Document15 pagesJPAM Vol 15 Issue1 P 285-299Jefri Nur HidayatNo ratings yet
- Paper 161Document7 pagesPaper 161Bilal SarwarNo ratings yet
- Molecules 27 01561 v2Document21 pagesMolecules 27 01561 v2sammedj2008No ratings yet
- Antibacterial Mechanism of Lactic Acid On Physiological andDocument6 pagesAntibacterial Mechanism of Lactic Acid On Physiological andlox standardNo ratings yet
- Antioxidants 12 00362 v2Document4 pagesAntioxidants 12 00362 v2Paskah DianNo ratings yet
- De Lucca 2011Document9 pagesDe Lucca 2011khadijaNo ratings yet
- Aquaculture: Bo Liu, Xianping Ge, Yanhui He, Jun Xie, Pao Xu, Yijin He, Qunlan Zhou, Liangkun Pan, Ruli ChenDocument7 pagesAquaculture: Bo Liu, Xianping Ge, Yanhui He, Jun Xie, Pao Xu, Yijin He, Qunlan Zhou, Liangkun Pan, Ruli ChenAndi Mahifa 93No ratings yet
- Original Article Antimicrobial and Antioxidant Activity of Crude Onion (Allium Cepa, L.) ExtractsDocument7 pagesOriginal Article Antimicrobial and Antioxidant Activity of Crude Onion (Allium Cepa, L.) ExtractsWidya LestariNo ratings yet
- Cytotoxic, Antibacterial, and Antioxidant Activities of The Leaf Extract of Sinningia BullataDocument18 pagesCytotoxic, Antibacterial, and Antioxidant Activities of The Leaf Extract of Sinningia BullataRodrigo ArrudaNo ratings yet
- Herbal Medicine and Parasitic Diseases: June 2011Document13 pagesHerbal Medicine and Parasitic Diseases: June 2011Sofia marisa fernandesNo ratings yet
- Antioxidant and Antibacterial Activities of Polyphenols From Ethnomedicinal Plants of Burkina FasoDocument6 pagesAntioxidant and Antibacterial Activities of Polyphenols From Ethnomedicinal Plants of Burkina Fasokaori_lawlietNo ratings yet
- Mushrooms and immunityDocument13 pagesMushrooms and immunityMartinaNo ratings yet
- Anti-Bacterial Activity of Garlic Extract AgainstDocument6 pagesAnti-Bacterial Activity of Garlic Extract AgainstSamantha Vhiel VicenteNo ratings yet
- Antioxidant and Antibacterial Activities of Thyme ExtractsDocument7 pagesAntioxidant and Antibacterial Activities of Thyme Extractsditya renaNo ratings yet
- Algal Bioactive Diversities Against Pathogenic MicrobesDocument8 pagesAlgal Bioactive Diversities Against Pathogenic Microbesmy.dear.sirNo ratings yet
- Molecules: Neuroprotective and Antioxidant Enhancing Properties of Selective Equisetum ExtractsDocument18 pagesMolecules: Neuroprotective and Antioxidant Enhancing Properties of Selective Equisetum ExtractsJoão FrancoNo ratings yet
- Antioxidant and Cytotoxic Activities of Centella Asiatica (L) UrbDocument9 pagesAntioxidant and Cytotoxic Activities of Centella Asiatica (L) UrbUwais AlNo ratings yet
- Potential of Plant Polyphenols To Combat Oxidative Stress and Inflammatory Processes in Farm Animals PDFDocument24 pagesPotential of Plant Polyphenols To Combat Oxidative Stress and Inflammatory Processes in Farm Animals PDFRafael SoaresNo ratings yet
- El GundousDocument19 pagesEl GundousANNE LOUISE FALCAO PASCOALNo ratings yet
- Molecules 26 04960Document21 pagesMolecules 26 04960ArielIzaSuquilloNo ratings yet
- Research Project 2Document4 pagesResearch Project 2valerybikobo588No ratings yet
- Comprehensive Review of Antimicrobial Activities of Plant FlavonoidsDocument32 pagesComprehensive Review of Antimicrobial Activities of Plant FlavonoidshusnulNo ratings yet
- Potential of Plant Polyphenols To Combat Oxidative Stress Andinflammatory Processes in Farm AnimalsDocument24 pagesPotential of Plant Polyphenols To Combat Oxidative Stress Andinflammatory Processes in Farm AnimalsRafael SoaresNo ratings yet
- Antibiotics: Research Progress On Antibacterial Activities and Mechanisms of Natural Alkaloids: A ReviewDocument30 pagesAntibiotics: Research Progress On Antibacterial Activities and Mechanisms of Natural Alkaloids: A Reviewriani.tiaraNo ratings yet
- s13659-023-00416-wDocument21 pagess13659-023-00416-wacheikhrouhouNo ratings yet
- Template Baru Addawaa 4.1Document5 pagesTemplate Baru Addawaa 4.1BIMA SATRIANo ratings yet
- Cell Growth Inhibitory Potential of Craterellus Cornucopioides (L.) Pers. Together With Antioxidant and Antimicrobial PropertiesDocument5 pagesCell Growth Inhibitory Potential of Craterellus Cornucopioides (L.) Pers. Together With Antioxidant and Antimicrobial Propertiesela.sofiaNo ratings yet
- Poultry Pathogen Antibacterial Activity of T. potatoriaDocument35 pagesPoultry Pathogen Antibacterial Activity of T. potatoriadavid ojNo ratings yet
- Konozy 2022 Bio Pharmacotherap Review Lectins As Antimicrobial AgentsDocument13 pagesKonozy 2022 Bio Pharmacotherap Review Lectins As Antimicrobial AgentsPatriciaNo ratings yet
- Lichen Secondary Metabolites As Potential Antibiotic Agents: 3.1 Needing For New Antibiotics From NatureDocument24 pagesLichen Secondary Metabolites As Potential Antibiotic Agents: 3.1 Needing For New Antibiotics From NatureIrfaan SalzabilNo ratings yet
- Molecules 28 02202Document16 pagesMolecules 28 02202Sandra Aliaga MorNo ratings yet
- Plantas Medicinales ColombianasDocument8 pagesPlantas Medicinales ColombianasBrigitte LilianaNo ratings yet
- Copy3 PDFDocument8 pagesCopy3 PDFPutri NafrianiiNo ratings yet
- Endophytes From Ethno Pharmacological Plants Sou - 2019 - Biocatalysis and AgriDocument17 pagesEndophytes From Ethno Pharmacological Plants Sou - 2019 - Biocatalysis and AgriMaria BatoolNo ratings yet
- Statia 20Document9 pagesStatia 20nguyen minh hienNo ratings yet
- Edible Plants As A Source of Antitubercular AgentsDocument7 pagesEdible Plants As A Source of Antitubercular AgentsRavi P ShaliwalNo ratings yet
- Yuvraj BiologyDocument19 pagesYuvraj BiologyAnkit KashyapNo ratings yet
- Antioxidant and Antimicrobial Activities of Oyster Mushroom: January 2017Document7 pagesAntioxidant and Antimicrobial Activities of Oyster Mushroom: January 2017AKNo ratings yet
- In Vitro Antioxidant Properties of Polysaccharides From Armillaria Mellea in Batch FermentationDocument10 pagesIn Vitro Antioxidant Properties of Polysaccharides From Armillaria Mellea in Batch FermentationJyoti PanchalNo ratings yet
- Sharma 2021Document14 pagesSharma 2021fibanezNo ratings yet
- Winska (2019) Essential Oils As Antimicrobial Agents-Myth or Real Alternative?Document21 pagesWinska (2019) Essential Oils As Antimicrobial Agents-Myth or Real Alternative?Phuong ThaoNo ratings yet
- Comparative Study of The Antibiotic Potency of NaturalDocument15 pagesComparative Study of The Antibiotic Potency of NaturalHuy NguyenNo ratings yet
- Screening of Antibacterial and Antioxidant Activities of Leaves ofDocument4 pagesScreening of Antibacterial and Antioxidant Activities of Leaves ofTiên Đỗ Ngọc ThúyNo ratings yet
- Klaus 2021Document14 pagesKlaus 2021Juan QuitianNo ratings yet
- 58-Article Text-269-1-10-20200406Document5 pages58-Article Text-269-1-10-20200406New CertificateNo ratings yet
- Antioxidants 10 00330 v2Document36 pagesAntioxidants 10 00330 v2StreetiiNo ratings yet
- Antimicrobial Activity Against Celtis AustralisDocument37 pagesAntimicrobial Activity Against Celtis AustralisarunNo ratings yet
- Metabolic Inhibitors V1: A Comprehensive TreatiseFrom EverandMetabolic Inhibitors V1: A Comprehensive TreatiseR HochsterNo ratings yet
- Element Content Botanical Composition and Nutritional Characteristics of NaturalDocument6 pagesElement Content Botanical Composition and Nutritional Characteristics of NaturalAytekin GaribliNo ratings yet
- Medicago Sativa Extractum Summary Report Committee Veterinary Medicinal Products - enDocument2 pagesMedicago Sativa Extractum Summary Report Committee Veterinary Medicinal Products - enAytekin GaribliNo ratings yet
- EP in Skin Care Video TranscriptDocument12 pagesEP in Skin Care Video TranscriptAytekin GaribliNo ratings yet
- Ijms 18 00512Document18 pagesIjms 18 00512Aytekin GaribliNo ratings yet
- 10 31195-Ejejfs 475286-603456Document4 pages10 31195-Ejejfs 475286-603456Aytekin GaribliNo ratings yet
- Trace Elements Analysis by Icpoes After Microwave Digestion of M 2014Document5 pagesTrace Elements Analysis by Icpoes After Microwave Digestion of M 2014Aytekin GaribliNo ratings yet
- Valerie Thompson - The Picture Book of A Day in Your Garden-Valerie Thompson Design (2021)Document43 pagesValerie Thompson - The Picture Book of A Day in Your Garden-Valerie Thompson Design (2021)Aytekin GaribliNo ratings yet
- View Free ArticleDocument9 pagesView Free ArticleAytekin GaribliNo ratings yet
- Nutritional Support For Immune Health: Sponsored byDocument22 pagesNutritional Support For Immune Health: Sponsored byAytekin GaribliNo ratings yet
- Lipotoxicidad Por Ceramidas CayetanoDocument13 pagesLipotoxicidad Por Ceramidas CayetanoLuciana Taco MendozaNo ratings yet
- Paracetamol 9Document6 pagesParacetamol 9danielafarmacie_1617No ratings yet
- Akp Important Guidelines 22 Guidlines Brief FinalDocument55 pagesAkp Important Guidelines 22 Guidlines Brief FinalRaheelNo ratings yet
- PharmacognRes 15 1 26Document16 pagesPharmacognRes 15 1 26Quyên ĐàoNo ratings yet
- Trends of Articles Published in National Journal of Physiology, Pharmacy and Pharmacology (NJPPP)Document7 pagesTrends of Articles Published in National Journal of Physiology, Pharmacy and Pharmacology (NJPPP)Amit ShahNo ratings yet
- 2020-2021 - Syllabus - English For Pharmacy PDFDocument80 pages2020-2021 - Syllabus - English For Pharmacy PDFElla ElaNo ratings yet
- Respiratory AgentDocument2 pagesRespiratory AgentAnthony LopezNo ratings yet
- Traditional Chinese Medicine Treatment of COVID-19Document2 pagesTraditional Chinese Medicine Treatment of COVID-19srinivas rao rayuduNo ratings yet
- MDS3 Ch06 Legislation Nov2011Document21 pagesMDS3 Ch06 Legislation Nov2011Andre SamsungNo ratings yet
- R. Basson. Human Sex-Response Cycles 2001Document11 pagesR. Basson. Human Sex-Response Cycles 2001Sylwia BarakehNo ratings yet
- Antidotes, Anticholinergics and Antibiotics: Amrit Pokhrel PharmacistDocument21 pagesAntidotes, Anticholinergics and Antibiotics: Amrit Pokhrel PharmacistheenamaharjanNo ratings yet
- Cytokine Release SyndromeDocument4 pagesCytokine Release SyndromeJuliandri JNo ratings yet
- Coronavirus Disease 2019 (COVID-19) - Cancer Screening, Diagnosis, Treatment, and Posttreatment Surveillance in Uninfected Patients During The Pandemic - UpToDateDocument55 pagesCoronavirus Disease 2019 (COVID-19) - Cancer Screening, Diagnosis, Treatment, and Posttreatment Surveillance in Uninfected Patients During The Pandemic - UpToDateTatiana Valencia CastañoNo ratings yet
- Drug Study RivastigmineDocument6 pagesDrug Study RivastigmineRaijenne VersolaNo ratings yet
- PT. BANGKIT ANUGERAH BERSAMA Product Price List Feb 2023Document11 pagesPT. BANGKIT ANUGERAH BERSAMA Product Price List Feb 2023dewi lestariNo ratings yet
- Cancer Pain Management-Part I: General PrinciplesDocument7 pagesCancer Pain Management-Part I: General PrinciplesNoviTrianaNo ratings yet
- Biosimilar Current Status in India: Review ArticleDocument4 pagesBiosimilar Current Status in India: Review ArticlebhupiscribdNo ratings yet
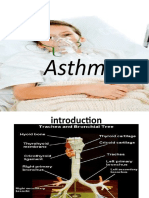
- Asthma (د فاكر القباطي)Document22 pagesAsthma (د فاكر القباطي)Aiman Al-SharabiNo ratings yet
- HSS - Summer 2020 - Syllabus - ANTH 1667Document7 pagesHSS - Summer 2020 - Syllabus - ANTH 1667Bruce JobseNo ratings yet
- A Review On Medicinal Uses of Sangu (Conch)Document4 pagesA Review On Medicinal Uses of Sangu (Conch)jmanuel108yahoo.co.ukNo ratings yet
- UPPER GASTROINTESTINAL BLEEDING (UGIB) MANAGEMENTDocument47 pagesUPPER GASTROINTESTINAL BLEEDING (UGIB) MANAGEMENTcopperNo ratings yet
- Overview of Complex Generics Regulatory Perspective On BioequivalenceDocument41 pagesOverview of Complex Generics Regulatory Perspective On BioequivalenceshrikantNo ratings yet
- Tinplate Qtn-2010-11 1Document51 pagesTinplate Qtn-2010-11 1Aditya SinghNo ratings yet
- Endoscopic Evaluation of Aceclofenac Gastrointestinal SafetyDocument11 pagesEndoscopic Evaluation of Aceclofenac Gastrointestinal SafetyMartin MoranNo ratings yet
- Nutritional Benefits in Cheese: Palanivel Ganesan Youn-Ho HongDocument24 pagesNutritional Benefits in Cheese: Palanivel Ganesan Youn-Ho HongasdfNo ratings yet
- Inhibitori, Inhibitorii Cresc: Substrat SubstraturileDocument4 pagesInhibitori, Inhibitorii Cresc: Substrat SubstraturileAlina CiugureanuNo ratings yet