You might also like
- Melo Et Al 2022 Sciadonus AlphacrucisDocument8 pagesMelo Et Al 2022 Sciadonus AlphacrucisMarcelo Roberto Souto de MeloNo ratings yet
- 22 Cormoran ImperialDocument8 pages22 Cormoran ImperialDavid Carmona LagosNo ratings yet
- Society For The Study of Amphibians and ReptilesDocument5 pagesSociety For The Study of Amphibians and ReptilesLĩnh NguyễnNo ratings yet
- Martins& Tavares - 2018aDocument6 pagesMartins& Tavares - 2018aLuciana MartinsNo ratings yet
- 02 First Record of Dolops StriataDocument4 pages02 First Record of Dolops StriataauzangatejbNo ratings yet
- Acuña 2013 First Record of Three Shark SpeciesDocument6 pagesAcuña 2013 First Record of Three Shark SpeciesHarry JonitzNo ratings yet
- Adaptive Radiation Versus Intraspeci®c Differentiation: Morphological Variation in Caribbean Anolis LizardsDocument6 pagesAdaptive Radiation Versus Intraspeci®c Differentiation: Morphological Variation in Caribbean Anolis LizardsMicaela GonellaNo ratings yet
- Aegla 2014Document14 pagesAegla 2014Ana VerdiNo ratings yet
- Thefossilrecordof AntarcticlandmammalsDocument20 pagesThefossilrecordof AntarcticlandmammalsAryan DubeyNo ratings yet
- Schwartz (1970) - Sphaerodactylus Notatus PDFDocument2 pagesSchwartz (1970) - Sphaerodactylus Notatus PDFKlaus FischerNo ratings yet
- Cunha Et AlDocument9 pagesCunha Et Alapi-3828346No ratings yet
- Bothrops Germanoi ARTIGODocument26 pagesBothrops Germanoi ARTIGOLigia AmorimNo ratings yet
- The Cardiidae Mollusca Bivalvia of TropiDocument90 pagesThe Cardiidae Mollusca Bivalvia of TropimuricidaeNo ratings yet
- tmpB44E TMPDocument5 pagestmpB44E TMPFrontiersNo ratings yet
- Amador Et Al 2017Document6 pagesAmador Et Al 2017Camila RodríguezNo ratings yet
- Equatoriana and Bolitoglossa Biseriata: Range: Notes On Geographic DistributionDocument4 pagesEquatoriana and Bolitoglossa Biseriata: Range: Notes On Geographic DistributionSilvia RosasNo ratings yet
- RossiSantos2009 CetaceanInteractionsDocument7 pagesRossiSantos2009 CetaceanInteractionsjenn.aereaNo ratings yet
- CheckList Article 18619 en 1Document3 pagesCheckList Article 18619 en 1kkfrita666No ratings yet
- cAPOlongo 2003Document18 pagescAPOlongo 2003Manuel IturriagaNo ratings yet
- Culebras Etal 2023Document4 pagesCulebras Etal 2023s6amplewNo ratings yet
- Check List: New Locality Records of Rhagomys Longilingua Luna & Patterson, 2003 (Rodentia: Cricetidae) in PeruDocument7 pagesCheck List: New Locality Records of Rhagomys Longilingua Luna & Patterson, 2003 (Rodentia: Cricetidae) in PeruMiluska R. SànchezNo ratings yet
- Phylogeographic Analysis of The Brown Alga Cutleria Multi Da (Tilopteridales, Phaeophyceae) Suggests A Complicated Introduction HistoryDocument8 pagesPhylogeographic Analysis of The Brown Alga Cutleria Multi Da (Tilopteridales, Phaeophyceae) Suggests A Complicated Introduction HistorySigfrido QuiñonesNo ratings yet
- 036 Chiasmocleis in The Atlantic Rain Forest CruzCaramaschiIzecksohn 1997 PDFDocument24 pages036 Chiasmocleis in The Atlantic Rain Forest CruzCaramaschiIzecksohn 1997 PDFAna Karolina MorenoNo ratings yet
- Neotropical Cervidology - 12Document13 pagesNeotropical Cervidology - 12Claudia DelafloresNo ratings yet
- On A New Genus and Species of Hemicytheridae (Ostracoda, Crustacea) From The Southern Brazilian CoastDocument7 pagesOn A New Genus and Species of Hemicytheridae (Ostracoda, Crustacea) From The Southern Brazilian Coastjoão carlos CoimbraNo ratings yet
- Baldo D., Et Al. 2008. Notes On Geographic Distribution. Reptilia, Gekkonidae, Hemidactylus Mabouia, Tarentola Mauritanica. Distribution Extension and Anthropogenic Dispersal.Document5 pagesBaldo D., Et Al. 2008. Notes On Geographic Distribution. Reptilia, Gekkonidae, Hemidactylus Mabouia, Tarentola Mauritanica. Distribution Extension and Anthropogenic Dispersal.leo836No ratings yet
- Copepoda Mesocyclops Ogunnus Matsumura e Silva 2002Document6 pagesCopepoda Mesocyclops Ogunnus Matsumura e Silva 2002Higor LessaNo ratings yet
- A New Species of From The Juan Ferna Ndez Islands: Eupithecia (Lepidoptera: Geometridae)Document7 pagesA New Species of From The Juan Ferna Ndez Islands: Eupithecia (Lepidoptera: Geometridae)Héctor Ibarra VidalNo ratings yet
- Amorphinopsis (Halichondrida: Demospongiae) From The Atlantic Ocean, With The Description of A New SpeciesDocument7 pagesAmorphinopsis (Halichondrida: Demospongiae) From The Atlantic Ocean, With The Description of A New SpeciesMara Paola DurangoNo ratings yet
- Original Article /artículo OriginalDocument18 pagesOriginal Article /artículo OriginalFreddy ASNo ratings yet
- Inshore Records of The Striped Dolphin S PDFDocument8 pagesInshore Records of The Striped Dolphin S PDFMary Clau Avila PeltrocheNo ratings yet
- Additional Anuran and Saurian Records For Phuket, South ThailandDocument5 pagesAdditional Anuran and Saurian Records For Phuket, South ThailandTeravut SuwansawaipholNo ratings yet
- Discovery of Extant (Chiroptera: Natalidae) in Cuba: Natalus MajorDocument10 pagesDiscovery of Extant (Chiroptera: Natalidae) in Cuba: Natalus Majormanuelperez_diezNo ratings yet
- Elefante Marino, MigracionesDocument7 pagesElefante Marino, MigracionesmayteNo ratings yet
- On A Small Collection of Bats (Chiroptera) From Western Sabah (North Borneo, East Malaysia)Document32 pagesOn A Small Collection of Bats (Chiroptera) From Western Sabah (North Borneo, East Malaysia)rabiatulfajriNo ratings yet
- Fosil de Tortuga en La HispaniolaDocument29 pagesFosil de Tortuga en La HispaniolaFedericoFerminNo ratings yet
- Phylogeography of The Bothrops JararacaDocument14 pagesPhylogeography of The Bothrops JararacaLigia AmorimNo ratings yet
- Taxonomy, Distribution, and Notes On The Termites (Isoptera: Kalotermitidae, Rhinotermitidae, Termitidae) of Puerto Rico and The U.S. Virgin IslandsDocument21 pagesTaxonomy, Distribution, and Notes On The Termites (Isoptera: Kalotermitidae, Rhinotermitidae, Termitidae) of Puerto Rico and The U.S. Virgin IslandsHanifah HerliniNo ratings yet
- Neo Tropical 12Document14 pagesNeo Tropical 12gabrielwerneckNo ratings yet
- Schwartz (1973) - Sphaerodactylus PDFDocument2 pagesSchwartz (1973) - Sphaerodactylus PDFKlaus FischerNo ratings yet
- Zootaxa: Sparisoma Tuiupiranga, A New Species of Parrotfish (PerciformesDocument14 pagesZootaxa: Sparisoma Tuiupiranga, A New Species of Parrotfish (PerciformesCarlos CavalcanteNo ratings yet
- Artículo 45 1 21 PDFDocument6 pagesArtículo 45 1 21 PDFLUISNo ratings yet
- Notas: New Locality Record of P (Squamata, Dipsadidae) in ArgentinaDocument4 pagesNotas: New Locality Record of P (Squamata, Dipsadidae) in ArgentinaClint AnthonyNo ratings yet
- Munilla Tomas & Soler Anna 2008Document13 pagesMunilla Tomas & Soler Anna 2008Enndy Jose Gonzalez ParejoNo ratings yet
- Animales ZoooMegaDocument19 pagesAnimales ZoooMegaClaudia TRNo ratings yet
- Introd Uzi DasDocument15 pagesIntrod Uzi DasDaiane MororóNo ratings yet
- Journal of South American Earth Sciences: A B C ADocument7 pagesJournal of South American Earth Sciences: A B C Amarburg 1967No ratings yet
- Genero MacrobrachiumDocument20 pagesGenero MacrobrachiumAna ParraNo ratings yet
- Amphipod Newsletter-25 2003 PDFDocument38 pagesAmphipod Newsletter-25 2003 PDFTheresa Marie ErmejeNo ratings yet
- AABC V83n1a20 PepesuchusDocument11 pagesAABC V83n1a20 PepesuchusKanekoNo ratings yet
- Bats of BarbadosDocument27 pagesBats of BarbadosJohnny BravoNo ratings yet
- 2002 Oreailurus Jacobita Description Comparision Felines Altiplano - GarciaPereaDocument15 pages2002 Oreailurus Jacobita Description Comparision Felines Altiplano - GarciaPereaLucía HNo ratings yet
- BothropsDocument3 pagesBothropsYostin AñinoNo ratings yet
- First Record of The Genus Fredilocarcinus Pretzmann, 1978 (Crustacea: Decapoda: Trichodactylidae) To ColombiaDocument2 pagesFirst Record of The Genus Fredilocarcinus Pretzmann, 1978 (Crustacea: Decapoda: Trichodactylidae) To ColombiaOphiophagusHannahNo ratings yet
- A New Small Short-Snouted Dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) From The Paleocene of Northeastern ColombiaDocument25 pagesA New Small Short-Snouted Dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) From The Paleocene of Northeastern ColombiaSERGIO MAYORGANo ratings yet
- Ma 2006 J Euk Micro Epistylis Onblue CrabDocument11 pagesMa 2006 J Euk Micro Epistylis Onblue CrabRIZKI NANDANo ratings yet
- Jaguarundi Herpailurus Yagoaroundi Notes in The Caribbean Lowlands of Belize and Guatemala Mccarthy 1992Document5 pagesJaguarundi Herpailurus Yagoaroundi Notes in The Caribbean Lowlands of Belize and Guatemala Mccarthy 1992María José TorallaNo ratings yet
- Tadpole, Oophagy, Advertisement Call, and Geographic Distribution of Aparasphenodon Arapapa Pimenta, Napoli and Haddad 2009 (Anura, Hylidae)Document6 pagesTadpole, Oophagy, Advertisement Call, and Geographic Distribution of Aparasphenodon Arapapa Pimenta, Napoli and Haddad 2009 (Anura, Hylidae)Guilherme SousaNo ratings yet
- Amarga-And-Fornesa 2020 LeptocyclopodiaDocument7 pagesAmarga-And-Fornesa 2020 LeptocyclopodiarabiatulfajriNo ratings yet
- 2021 The Cost of Gold Mercury ContaminationDocument24 pages2021 The Cost of Gold Mercury ContaminationcomandocrawNo ratings yet
- Foraging Theory and Prey Size-Predator Size Relations in SnakesDocument31 pagesForaging Theory and Prey Size-Predator Size Relations in SnakescomandocrawNo ratings yet
- Sosa Etal 2019 Herp ReviewDocument3 pagesSosa Etal 2019 Herp ReviewcomandocrawNo ratings yet
- Schalk Etal 2018 HRDocument6 pagesSchalk Etal 2018 HRcomandocrawNo ratings yet
- 3-MS Examples PDFDocument59 pages3-MS Examples PDFlexuanminhk54lochoaNo ratings yet
- UltraSonic Processors - CatalogDocument2 pagesUltraSonic Processors - Catalogdéborah_rosalesNo ratings yet
- Ovulation CalanderDocument2 pagesOvulation CalanderBN NGNo ratings yet
- Sociology Handout 5Document13 pagesSociology Handout 5Carzo Aggy MugyNo ratings yet
- Clave Genero Dermanura (Otros) PDFDocument520 pagesClave Genero Dermanura (Otros) PDFJuan D Garcia Peluffo100% (1)
- NutritionDocument5 pagesNutritionk,srikanthNo ratings yet
- Trabalho em Saúde e A Implantação Do Acolhimento Na Atenção Primária À Saúde - Afeto, Empatia Ou AlteridadeDocument13 pagesTrabalho em Saúde e A Implantação Do Acolhimento Na Atenção Primária À Saúde - Afeto, Empatia Ou AlteridadeAmérico PastorNo ratings yet
- Circulatory SystemDocument16 pagesCirculatory SystemHarinder KaurNo ratings yet
- Blood Doping PPT PresentationDocument26 pagesBlood Doping PPT Presentationee4254100% (1)
- PDFDocument570 pagesPDFEshaal FatimaNo ratings yet
- Jellyfish Research PaperDocument6 pagesJellyfish Research Papergzrvpcvnd100% (1)
- Proposed Pre Final-1 Time Table With Weightage.. MPC, Bipc&Civils-AP&TsDocument17 pagesProposed Pre Final-1 Time Table With Weightage.. MPC, Bipc&Civils-AP&TsAbhiram muddanaNo ratings yet
- Staar Eoc 2016test Bio F 7Document39 pagesStaar Eoc 2016test Bio F 7api-293216402No ratings yet
- Pharyngeal ArchesDocument35 pagesPharyngeal ArchesconstantinilieNo ratings yet
- The Effect of Temperature On The Reaction Time of The Enzyme Peroxidase - Lab ReportDocument5 pagesThe Effect of Temperature On The Reaction Time of The Enzyme Peroxidase - Lab ReportAnonymous aZEERFhvNo ratings yet
- FTS - 11 (Code-A) - 24-04-2020 PDFDocument16 pagesFTS - 11 (Code-A) - 24-04-2020 PDFkavyareddyNo ratings yet
- Contoh Arsitektur SimbioisisDocument12 pagesContoh Arsitektur SimbioisisDeni AtahillahNo ratings yet
- Cell and Tissue Culture: Model Cells in Medicine - Stem Cells - Tissue EngineeringDocument44 pagesCell and Tissue Culture: Model Cells in Medicine - Stem Cells - Tissue EngineeringCrasilia Yanti PadangNo ratings yet
- Using STADEN For Sequence AssemblyDocument5 pagesUsing STADEN For Sequence AssemblyChristen Rune StensvoldNo ratings yet
- Approved LaboratoriesDocument22 pagesApproved LaboratoriesPaul CNo ratings yet
- 3 - Plasma Membrane and Transport MechanismsDocument67 pages3 - Plasma Membrane and Transport MechanismsThom PaglinawanNo ratings yet
- ART - Spectrophotometric Method For Determining GA in ..Document4 pagesART - Spectrophotometric Method For Determining GA in ..Azahara Linares100% (2)
- The Impact of Your Internal Terrain On Your FasciaDocument16 pagesThe Impact of Your Internal Terrain On Your Fasciazzip1961100% (1)
- Gender and PsychotherapyDocument12 pagesGender and PsychotherapyEhh EnNaNo ratings yet
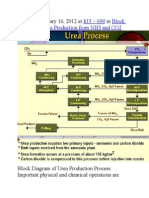
- Published January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2Document9 pagesPublished January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2himanshuchawla654No ratings yet
- Tushar FinalDocument29 pagesTushar FinalRaj Prixit RathoreNo ratings yet
- OB Ultrasound Report Template 2Document1 pageOB Ultrasound Report Template 2PriyankaNo ratings yet
- Test On Gmos GM Foods Cloning b1b2 Grammar Drills Information Gap Activities Reading 96207Document4 pagesTest On Gmos GM Foods Cloning b1b2 Grammar Drills Information Gap Activities Reading 96207Thu Quynh NguyenNo ratings yet
- Biosensors For Environmental ApplicationsDocument15 pagesBiosensors For Environmental ApplicationsBenjamin HonorioNo ratings yet
- Selina Solutions For Class 9 Biology Chapter 2 Cell The Unit of LifeDocument15 pagesSelina Solutions For Class 9 Biology Chapter 2 Cell The Unit of LifePrashun PriyadarshiNo ratings yet