You might also like
- The Bioavailability of Dietary CalciumDocument18 pagesThe Bioavailability of Dietary CalciumtvmedicineNo ratings yet
- Pregnancy and Lactation in Adolescence: Possible Implications For Calcium Metabolism and Bone MassDocument12 pagesPregnancy and Lactation in Adolescence: Possible Implications For Calcium Metabolism and Bone MassDoggoboy WolfmannNo ratings yet
- Document YambarinesDocument5 pagesDocument YambarinespetrtesssgNo ratings yet
- Department of Periodontics and Implantology: Calcium MetabolismDocument63 pagesDepartment of Periodontics and Implantology: Calcium Metabolismrasagna reddy100% (1)
- Calcium - Hahn Mcnutt ReddmannDocument15 pagesCalcium - Hahn Mcnutt Reddmannapi-332181335No ratings yet
- Calcium Supplements An OverviewDocument7 pagesCalcium Supplements An OverviewnganhaNo ratings yet
- Calcium MetabolismDocument80 pagesCalcium MetabolismaayushNo ratings yet
- The Bioavailability of Dietary Calcium PDFDocument18 pagesThe Bioavailability of Dietary Calcium PDFStev D'BulletNo ratings yet
- RBTI Insights For Super Healthy BonesDocument4 pagesRBTI Insights For Super Healthy Bones786lailaNo ratings yet
- Fasting With Calcium Ion Therapy 1Document10 pagesFasting With Calcium Ion Therapy 1tjwtzkpnz8No ratings yet
- Dietary Needs Forhealthy BonesDocument7 pagesDietary Needs Forhealthy BonesAlli NdahuraNo ratings yet
- Corrected CHAPTER I - SubhathraDocument6 pagesCorrected CHAPTER I - SubhathraGnana PrakashNo ratings yet
- Nutrional Aspects of Calcium AbsorptionDocument4 pagesNutrional Aspects of Calcium Absorptiondian aristiaNo ratings yet
- Movilidad MadridDocument9 pagesMovilidad Madridubiktrash1492No ratings yet
- Clinical Application of Calcium Modeling in Patients With Chronic Kidney Disease.Document4 pagesClinical Application of Calcium Modeling in Patients With Chronic Kidney Disease.Kimberly SemenzaNo ratings yet
- MineralsDocument9 pagesMineralsAyesha AfzalNo ratings yet
- Ann. Din. Biochem.: Regulation of Calcium MetabolismDocument22 pagesAnn. Din. Biochem.: Regulation of Calcium MetabolismAzmi FarhadiNo ratings yet
- Osteo Book PdaDocument53 pagesOsteo Book PdaArpanpatelNo ratings yet
- American Academy of Pediatrics: PEDIATRICS Vol. 104 No. 5 November 1999Document8 pagesAmerican Academy of Pediatrics: PEDIATRICS Vol. 104 No. 5 November 1999AnnikaNo ratings yet
- 19 - Lecture #14 - Minerals - Part 1Document58 pages19 - Lecture #14 - Minerals - Part 1Alexandra AyneNo ratings yet
- Bone Formula Us PipDocument3 pagesBone Formula Us PipThanh VoNo ratings yet
- Health in Your Hands: By: Ch. Saqib Abbas 1Document15 pagesHealth in Your Hands: By: Ch. Saqib Abbas 1AdilMinhajNo ratings yet
- 10-Nutritional Considerations in Joint HealthDocument18 pages10-Nutritional Considerations in Joint Healthapi-3851239No ratings yet
- 783.full Calcium Bioav Absorption AllenDocument26 pages783.full Calcium Bioav Absorption Allenadrip234No ratings yet
- The Role of Diet in Osteoporosis Prevention and Management: Epidemiology and Pathophysiology (Ra Adler, Section Editor)Document7 pagesThe Role of Diet in Osteoporosis Prevention and Management: Epidemiology and Pathophysiology (Ra Adler, Section Editor)Jamaluddin HaikhahNo ratings yet
- Newsletter 2007 SpringdDocument4 pagesNewsletter 2007 Springd786lailaNo ratings yet
- The Data of Science: CreationDocument14 pagesThe Data of Science: CreationAbraham AbsalonNo ratings yet
- MagChlor PDFDocument6 pagesMagChlor PDFDonte WeberNo ratings yet
- Intake: Recommended Intakes of Bone-Related Nutrients For AdultsDocument1 pageIntake: Recommended Intakes of Bone-Related Nutrients For AdultsAnonymous 4j7k1q9HNo ratings yet
- AAP 2210 Nutrition and Feeding of Non-Ruminants - Last Part 22.11.2019Document33 pagesAAP 2210 Nutrition and Feeding of Non-Ruminants - Last Part 22.11.2019Kevin KahingaNo ratings yet
- CalciumDocument3 pagesCalciummoussa.orgNo ratings yet
- Pscalcium06 PDFDocument16 pagesPscalcium06 PDFObed Garcia EscarpulliNo ratings yet
- Unit-5 The Micronutrients-II MineralsDocument12 pagesUnit-5 The Micronutrients-II MineralsStanley WangchokNo ratings yet
- Na and OsteoDocument6 pagesNa and Osteop1kac4uNo ratings yet
- 6 - J PerinetalDocument6 pages6 - J PerinetaltaimoursultanNo ratings yet
- Nutrition and BoneDocument6 pagesNutrition and BonedionyessNo ratings yet
- Assignment ON: Ccs Haryana Agriculture University HisarDocument16 pagesAssignment ON: Ccs Haryana Agriculture University HisarVinod KumarNo ratings yet
- Calcium Metabolism PraveenDocument42 pagesCalcium Metabolism PraveenDr PraveenNo ratings yet
- Personal Nutrition 9th Edition Boyle Solutions ManualDocument32 pagesPersonal Nutrition 9th Edition Boyle Solutions Manualthoabangt69100% (26)
- Chapter 23: Osteoporosis of The SpineDocument15 pagesChapter 23: Osteoporosis of The SpineTraian UrsuNo ratings yet
- Musculoskeltal SystemDocument18 pagesMusculoskeltal Systemhy7tnNo ratings yet
- Calcium: Basic FunctionsDocument4 pagesCalcium: Basic FunctionsTriesha Mae GalangNo ratings yet
- Calcium - The Nutrition Source - Harvard T.H. Chan School of Public HealthDocument6 pagesCalcium - The Nutrition Source - Harvard T.H. Chan School of Public HealthSarah DanielNo ratings yet
- A System To Assess The Quality of Food Sources of Calcium: C. Alan Titchenal, Joannie DobbsDocument8 pagesA System To Assess The Quality of Food Sources of Calcium: C. Alan Titchenal, Joannie DobbsReZqie WaNa PradyPthaNo ratings yet
- The Importance of Minerals in The Health of HumansDocument11 pagesThe Importance of Minerals in The Health of Humanskrich navarroNo ratings yet
- CalciumDocument41 pagesCalciumIka Nindyas RanitadewiNo ratings yet
- Vitamins and Menerals To ReadDocument17 pagesVitamins and Menerals To ReadVivek VaishnavNo ratings yet
- Recommendations To Prevent and Control Iron Deficiency in The United StatesDocument28 pagesRecommendations To Prevent and Control Iron Deficiency in The United StatesAndiFitryanaNo ratings yet
- Caffeine Bone CalciumDocument8 pagesCaffeine Bone CalciumHerpika DianaNo ratings yet
- Ijem 23 387Document4 pagesIjem 23 387Ahsan KhanNo ratings yet
- CALCIUMDocument30 pagesCALCIUMMona Riza PagtabonanNo ratings yet
- Calcium-Magnesium Complex™ Nourish Bones and Regulate Growth7.35ozDocument3 pagesCalcium-Magnesium Complex™ Nourish Bones and Regulate Growth7.35oztipitakastudiesNo ratings yet
- Porose Gravidez e Puerperio 2021Document15 pagesPorose Gravidez e Puerperio 2021Ben-Hur AlbergariaNo ratings yet
- ABStracts Su Componenti EndosilanDocument14 pagesABStracts Su Componenti EndosilanRiccardo FlorisNo ratings yet
- Miapoly Calcium in AnimalDocument8 pagesMiapoly Calcium in AnimalDev jibreenNo ratings yet
- Intro To StarsDocument37 pagesIntro To StarsENo ratings yet
- Dassault Systems Academic CalenderDocument5 pagesDassault Systems Academic CalenderSarath KumarNo ratings yet
- BBC Learning English - 6 Minute English - Bitcoin - Digital Crypto-CurrencyDocument38 pagesBBC Learning English - 6 Minute English - Bitcoin - Digital Crypto-CurrencyMohamad GhafooryNo ratings yet
- Major06 QP DLP NEET2019 (Pmtcorner - In) PDFDocument40 pagesMajor06 QP DLP NEET2019 (Pmtcorner - In) PDFMegha HazarikaNo ratings yet
- LNMIIT Course Information Form: A. B. C. D. E. FDocument2 pagesLNMIIT Course Information Form: A. B. C. D. E. FAayush JainNo ratings yet
- EKOFLUID PRODUCT SHEET FILOIL 6000 210x297 EN WEBDocument4 pagesEKOFLUID PRODUCT SHEET FILOIL 6000 210x297 EN WEBjunaidi.036No ratings yet
- DiseasesDocument11 pagesDiseasesapi-307430346No ratings yet
- DSP UPG CU EngDocument1 pageDSP UPG CU EngArtur KwiatkowskiNo ratings yet
- Tea Board of India PDFDocument18 pagesTea Board of India PDFDebasish RazNo ratings yet
- John Deere CaseDocument2 pagesJohn Deere CaseAldo ReynaNo ratings yet
- Marking Scheme According To AIDocument2 pagesMarking Scheme According To AIAbdul RehmanNo ratings yet
- 2006 317Document13 pages2006 317LonginoNo ratings yet
- Aeroacoustic Optimization of Wind Turbine Airfoils by Combining Thermographic and Acoustic Measurement DataDocument4 pagesAeroacoustic Optimization of Wind Turbine Airfoils by Combining Thermographic and Acoustic Measurement DatamoussaouiNo ratings yet
- Inspection and Test Plan (ITP) For Spherical Storage Tanks: Dehloran Olefin PlantDocument9 pagesInspection and Test Plan (ITP) For Spherical Storage Tanks: Dehloran Olefin PlantbahmanNo ratings yet
- 1250kva DG SetDocument61 pages1250kva DG SetAnagha Deb100% (1)
- James Bruce, One of Russian Tsar Peter The Great's Key Advisors (1669-1735)Document2 pagesJames Bruce, One of Russian Tsar Peter The Great's Key Advisors (1669-1735)Johanna Granville100% (1)
- Answerkey Precise ListeningDocument26 pagesAnswerkey Precise ListeningAn LeNo ratings yet
- Management From RamayanaDocument14 pagesManagement From Ramayanasaaket batchuNo ratings yet
- 2946 1308 00 Leroy Somer LSA47.2 Installation & MaintenanceDocument20 pages2946 1308 00 Leroy Somer LSA47.2 Installation & MaintenanceJORGE ARMANDO CARRASCO TICLLENo ratings yet
- HLTARO001 HLTAROO05 Student Assessment Booklet 1 1Document68 pagesHLTARO001 HLTAROO05 Student Assessment Booklet 1 1Amber PreetNo ratings yet
- TelemetryDocument8 pagesTelemetryKristin WiseNo ratings yet
- Instability of Slender Concrete Deep BeamDocument12 pagesInstability of Slender Concrete Deep BeamFrederick TanNo ratings yet
- Tahmina Ferdousy Jhumu: HND Btec Unit 15 Psychology For Health and Social CareDocument29 pagesTahmina Ferdousy Jhumu: HND Btec Unit 15 Psychology For Health and Social CareNabi BoxNo ratings yet
- 78EZDM Product Specifications Conector para Cable de 7 OctavosDocument4 pages78EZDM Product Specifications Conector para Cable de 7 OctavosEdwin Santiago QuispeNo ratings yet
- Organizational Behavior Exam 2 Practice QuestionsDocument1 pageOrganizational Behavior Exam 2 Practice QuestionsSydney EverettNo ratings yet
- Brand Loyalty & Competitive Analysis of Pankaj NamkeenDocument59 pagesBrand Loyalty & Competitive Analysis of Pankaj NamkeenBipin Bansal Agarwal100% (1)
- Āyāt Al-Aḥkām, Ayat Al-AhkamDocument6 pagesĀyāt Al-Aḥkām, Ayat Al-AhkamRasoul NamaziNo ratings yet
- English The Smiles and Tears of RasoolullahDocument130 pagesEnglish The Smiles and Tears of RasoolullahwildqafNo ratings yet
- Dharmakirti On Pratyaksa PDFDocument14 pagesDharmakirti On Pratyaksa PDFonlineyyk100% (1)
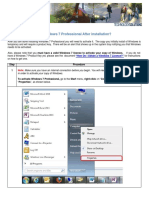
- Activate My Win 7 Pro Product KeyDocument4 pagesActivate My Win 7 Pro Product KeydoktauNo ratings yet