You might also like
- Omniverse PDFDocument429 pagesOmniverse PDFJorge Barios100% (12)
- Tract Continues Down Spinal Cord: Input Location Notes Lesion MedullaDocument10 pagesTract Continues Down Spinal Cord: Input Location Notes Lesion MedullaherethemindNo ratings yet
- Neuroanatomy Revision Notes PDFDocument7 pagesNeuroanatomy Revision Notes PDFTanya Tanu100% (1)
- Tracts of The Spinal CordDocument21 pagesTracts of The Spinal Corddr.a.k.gupta6924100% (10)
- Diploma in Occupational TherapyDocument47 pagesDiploma in Occupational TherapyAzmi Plus100% (2)
- The Midbrain and Important ConnectionsDocument118 pagesThe Midbrain and Important ConnectionsBaguma MichaelNo ratings yet
- Diploma in Occupational TherapyDocument21 pagesDiploma in Occupational TherapyAzmi Plus100% (1)
- Introduction and Organization of Nervous System: Presented by Dr. Amen FatimaDocument37 pagesIntroduction and Organization of Nervous System: Presented by Dr. Amen FatimaShanailNo ratings yet
- Edexcel Ial Biology Lab BookpdfDocument69 pagesEdexcel Ial Biology Lab Bookpdfbgtes12367% (6)
- Sex Determination and Sex Chromosomes: Lecture Presentations by Cindy Malone California State University, NorthridgeDocument53 pagesSex Determination and Sex Chromosomes: Lecture Presentations by Cindy Malone California State University, NorthridgeAudrey100% (1)
- Cerebellum TextDocument7 pagesCerebellum TextJean Pierre Chastre LuzaNo ratings yet
- Department of Anatomy College of Medicine University of Ibadan Neuroanatomy Dissection of The BrainDocument22 pagesDepartment of Anatomy College of Medicine University of Ibadan Neuroanatomy Dissection of The BrainAbdulateef AdebisiNo ratings yet
- Summary of Pathways 2020 5e and 6e MedNeuroDocument25 pagesSummary of Pathways 2020 5e and 6e MedNeuromaria fernandaNo ratings yet
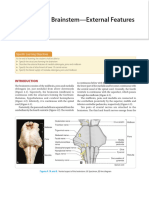
- Brainstem: External and Internal FeaturesDocument57 pagesBrainstem: External and Internal FeaturesJoaquin GuillermoNo ratings yet
- Cerebellum Gross Appearance of The Cerebellum: Posterior Lobe), Which Is TheDocument10 pagesCerebellum Gross Appearance of The Cerebellum: Posterior Lobe), Which Is TheRafu DestajoNo ratings yet
- Brain Stem FullDocument25 pagesBrain Stem FullCosplay AccountNo ratings yet
- Brainstem (Gross and Sectional Anatomy) : Internal Structure of Medulla-PonsDocument3 pagesBrainstem (Gross and Sectional Anatomy) : Internal Structure of Medulla-PonsPrajjwal GuptaNo ratings yet
- Dorsal Column-Medial Lemniscus Pathway - WikipediaDocument27 pagesDorsal Column-Medial Lemniscus Pathway - WikipediaCosplay AccountNo ratings yet
- 01 Neuroscience: Gross Anatomy of The BrainDocument5 pages01 Neuroscience: Gross Anatomy of The BrainAsh CansanayNo ratings yet
- Introduction & Organization of The Nervous System: Marlene Ricci Castillo, MD, FPCS, FpsgsDocument39 pagesIntroduction & Organization of The Nervous System: Marlene Ricci Castillo, MD, FPCS, FpsgsAfNo ratings yet
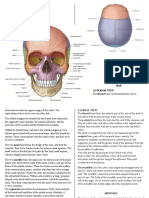
- Skull Anterior View: The Frontal Bone (Or Forehead Bone) CurvesDocument22 pagesSkull Anterior View: The Frontal Bone (Or Forehead Bone) CurvesFatima JamalNo ratings yet
- Brain StemDocument4 pagesBrain StemHeart Of Ayurveda Beatrice DNo ratings yet
- White Matter of The Cerebral HemispheresDocument50 pagesWhite Matter of The Cerebral HemispheresSandy MonirNo ratings yet
- NBSS CASE 6 - Cerebral Palsy and Mental RetardationDocument28 pagesNBSS CASE 6 - Cerebral Palsy and Mental RetardationbungadielaNo ratings yet
- Internal Structure of Pons: 1-Lower Level 2-Mid-Level: 3-Upper LevelDocument28 pagesInternal Structure of Pons: 1-Lower Level 2-Mid-Level: 3-Upper LevelsridharNo ratings yet
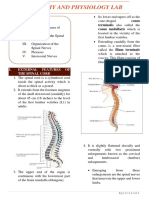
- Spinal CordDocument6 pagesSpinal Cordsangesh kNo ratings yet
- Brainstem: Medulla OblongataDocument130 pagesBrainstem: Medulla OblongataBhawana BhatodiyaNo ratings yet
- Aprimaion Angulo PontocerebelosoDocument64 pagesAprimaion Angulo PontocerebelosoLuis Carlos Avellaneda CurchoNo ratings yet
- Chapter 9 - BrainDocument9 pagesChapter 9 - Brainbussniesm05No ratings yet
- TA Lab 5 BrainDocument9 pagesTA Lab 5 BrainHajira NusretNo ratings yet
- Assignment#4 Clinical Neuro Sciences: TOPIC: Describe Briefly The Clinical Testing of All Cranial NervesDocument3 pagesAssignment#4 Clinical Neuro Sciences: TOPIC: Describe Briefly The Clinical Testing of All Cranial NervesAlina ShaukatNo ratings yet
- Chapter 3 - Structure of The Nervous SystemDocument8 pagesChapter 3 - Structure of The Nervous Systemmanilyn dacoNo ratings yet
- Neuroscience: Topic: MeningesDocument6 pagesNeuroscience: Topic: MeningesRoselle AbrazaldoNo ratings yet
- Glossary of Neuroanatomy TermsDocument10 pagesGlossary of Neuroanatomy Termsmohan730463No ratings yet
- Medulla OblongataDocument9 pagesMedulla OblongataExtra74 ExtraNo ratings yet
- Atlas of Microscopic Anatomy InterDocument29 pagesAtlas of Microscopic Anatomy InterIndraPerdanaNo ratings yet
- Modulation of Movement by The CerebellumDocument14 pagesModulation of Movement by The Cerebellumgiahan.lieuNo ratings yet
- Spinal CordDocument50 pagesSpinal CordReem 10No ratings yet
- CNS PathwaysDocument38 pagesCNS PathwaysVitor MartinsNo ratings yet
- Safari - Feb 21, 2024 at 12:17 PMDocument1 pageSafari - Feb 21, 2024 at 12:17 PMsyansyncNo ratings yet
- 6th Lecture - Physiology of The Brain Stem - CNS PhysiologyDocument13 pages6th Lecture - Physiology of The Brain Stem - CNS PhysiologyAnastasia SoareNo ratings yet
- Physio. Psych Structure-Of-The-Nervous-System-Fact-SheetDocument7 pagesPhysio. Psych Structure-Of-The-Nervous-System-Fact-SheetJustynmae JusainNo ratings yet
- Neuroanatomy, Brainstem - StatPearls - NCBI BookshelfDocument13 pagesNeuroanatomy, Brainstem - StatPearls - NCBI BookshelfKritika MalhotraNo ratings yet
- Brainstem Stroke SyndromesDocument43 pagesBrainstem Stroke SyndromesSarah Choo Qian RouNo ratings yet
- 48-Hour Chick Embryo: ST ND RDDocument29 pages48-Hour Chick Embryo: ST ND RDFerhaeeza KalayakanNo ratings yet
- CerebrumDocument2 pagesCerebrumjnthnrsjNo ratings yet
- Brain Stem: Rabbia Yousaf (Pharm-D) Rashid Latif College of PharmacyDocument30 pagesBrain Stem: Rabbia Yousaf (Pharm-D) Rashid Latif College of PharmacyShafaqat Ghani Shafaqat GhaniNo ratings yet
- Spinal CordDocument8 pagesSpinal CordCUESCANO, CRISTEL G. BSMT 2-ANo ratings yet
- Protection of The CNSDocument4 pagesProtection of The CNSGrayson JamesNo ratings yet
- The Medulla Oblongata - Internal Structure - Vasculature - TeachMeAnatomyDocument6 pagesThe Medulla Oblongata - Internal Structure - Vasculature - TeachMeAnatomyraihaana turawaNo ratings yet
- Midbrain - 2-18-07-2020Document26 pagesMidbrain - 2-18-07-2020Vinod SharmaNo ratings yet
- Gray's Anatomy - The Anatomical Basis of Clinical Practice - Chap 19 Doc HijosaDocument8 pagesGray's Anatomy - The Anatomical Basis of Clinical Practice - Chap 19 Doc Hijosatiburciamd27No ratings yet
- The Posterior Fossa VeinsDocument24 pagesThe Posterior Fossa Veinsbodeadumitru9261100% (2)
- Spinal Cord 1Document8 pagesSpinal Cord 1Alina PetreNo ratings yet
- MHSB Lab - Brainstem External FeaturesDocument6 pagesMHSB Lab - Brainstem External FeaturesVee ManahanNo ratings yet
- 16-Brainstem - Agdmd Oct 2022Document137 pages16-Brainstem - Agdmd Oct 2022vbh8222No ratings yet
- Topognosis of Facial Nerve LesionsDocument11 pagesTopognosis of Facial Nerve LesionsAudiologiaNo ratings yet
- Failure of Neuropores To Close Can Cause Neural Tube DefectsDocument6 pagesFailure of Neuropores To Close Can Cause Neural Tube DefectsMargo Roth SpiegelmanNo ratings yet
- Gross B 2.20. Superficial Structures and Posterior Triangle of The NeckDocument6 pagesGross B 2.20. Superficial Structures and Posterior Triangle of The NeckShaira Aquino VerzosaNo ratings yet
- Capitulo 4Document12 pagesCapitulo 4Carla MonteroNo ratings yet
- Embryology of The CNS 2022Document130 pagesEmbryology of The CNS 2022isicheipraise3No ratings yet
- DEVELOPMENT NERVOUS SYSTEM Brain Face PDFDocument15 pagesDEVELOPMENT NERVOUS SYSTEM Brain Face PDFChidera EmmanuelNo ratings yet
- Biology Cell NotesDocument2 pagesBiology Cell NotesayanNo ratings yet
- Sf2 2019 Grade 7 Year I HonestyDocument6 pagesSf2 2019 Grade 7 Year I HonestyDaniel ManuelNo ratings yet
- New Microsoft Office Word DocumentDocument5 pagesNew Microsoft Office Word DocumentPal AlexandraNo ratings yet
- Plant Collections From Bangladesh in The Herbarium at Shillong (Assam), India.Document26 pagesPlant Collections From Bangladesh in The Herbarium at Shillong (Assam), India.López LuisNo ratings yet
- 2023 05 14 540710v1 FullDocument17 pages2023 05 14 540710v1 Fullliukeyin.nccNo ratings yet
- (Izuru Yamamoto (Auth.), Izuru Yamamoto, John E. C PDFDocument299 pages(Izuru Yamamoto (Auth.), Izuru Yamamoto, John E. C PDFJuliana BarrosoNo ratings yet
- N9 แนวข้อสอบวิชาภาษาอังกฤษDocument30 pagesN9 แนวข้อสอบวิชาภาษาอังกฤษJO-JO17No ratings yet
- Soal Dan Pembahasan UN Bahasa Inggris SMA IPS 2009-2010Document15 pagesSoal Dan Pembahasan UN Bahasa Inggris SMA IPS 2009-2010Genius Edukasi50% (2)
- Gene Delivery ThesisDocument5 pagesGene Delivery ThesisHelpWritingPapersUK100% (2)
- DPM 55Document12 pagesDPM 55Sureshi BishtNo ratings yet
- Unit 3Document11 pagesUnit 3gachatoqaNo ratings yet
- Literature Review On Rice CultivationDocument7 pagesLiterature Review On Rice Cultivationfdnmffvkg100% (1)
- AmphibiansandreptilesDocument14 pagesAmphibiansandreptilesYeeyee AungNo ratings yet
- An In-Depth Look at Krukenberg TumorDocument6 pagesAn In-Depth Look at Krukenberg Tumor垂直马克No ratings yet
- Chapter 1 Introduction To EcologyDocument30 pagesChapter 1 Introduction To Ecologynurul syafiyah binti yusoffNo ratings yet
- Admission Notice For Gat-B Students 2020 - 2021Document2 pagesAdmission Notice For Gat-B Students 2020 - 2021Mohit JagtapNo ratings yet
- 5 DiscussionDocument5 pages5 DiscussionAnowar MasukNo ratings yet
- Sample A&P QuestionsDocument4 pagesSample A&P QuestionsAmala SmithNo ratings yet
- Arecaceae or Palmaceae (Palm Family) : Reported By: Kerlyn Kaye I. AsuncionDocument23 pagesArecaceae or Palmaceae (Palm Family) : Reported By: Kerlyn Kaye I. AsuncionNamae BacalNo ratings yet
- Indoor Air Quality and Student PerformanceDocument8 pagesIndoor Air Quality and Student Performancesunnews820No ratings yet
- MS-SCI-LS-Unit 1 - Chapter 3 - Cell Structure and Function PDFDocument38 pagesMS-SCI-LS-Unit 1 - Chapter 3 - Cell Structure and Function PDFNNo ratings yet
- Alphaview User GuideDocument10 pagesAlphaview User GuideMichael CaverlyNo ratings yet
- Newsletter: Plant Mutation BreedingDocument3 pagesNewsletter: Plant Mutation BreedingGabbyNo ratings yet
- Cat 31032631Document284 pagesCat 31032631semahegnyitayewNo ratings yet
- Project List-Biology ACADEMIC SESSION: - 2022-2023: CBSE AFFILIATION No.: - 1130522Document7 pagesProject List-Biology ACADEMIC SESSION: - 2022-2023: CBSE AFFILIATION No.: - 1130522Neehar GuitarNo ratings yet
- Wound CultureDocument10 pagesWound CultureIndah SusantiNo ratings yet
- Cholin BaseDocument5 pagesCholin Basex_aav3No ratings yet