You might also like
- Lipid Signaling and MetabolismFrom EverandLipid Signaling and MetabolismJames M. NtambiNo ratings yet
- EritrocitosYdefectos MetabolicosDocument10 pagesEritrocitosYdefectos Metabolicosandres herreraNo ratings yet
- Nucleic Acid FultonDocument21 pagesNucleic Acid FultonMicky Amekan100% (1)
- JTR2014 649502 PDFDocument7 pagesJTR2014 649502 PDFIlmanNo ratings yet
- Clinical Notes For BiochemistryDocument8 pagesClinical Notes For Biochemistrymoathreaq620No ratings yet
- Rac1/Pi3K/Pkb-Dependent Caspase-1 Activation Release Through Hmg-Coa Reductase Inhibition Induces Il-1Document12 pagesRac1/Pi3K/Pkb-Dependent Caspase-1 Activation Release Through Hmg-Coa Reductase Inhibition Induces Il-1Josse BouwhuisNo ratings yet
- CompoundsDocument6 pagesCompoundsnr6zqkdcdgNo ratings yet
- PKD BiochemistryDocument7 pagesPKD BiochemistryHyun Jae WonNo ratings yet
- Mechanism of Activation of The Cdk2Document3 pagesMechanism of Activation of The Cdk2MUHAMMAD ARIFFUDINNo ratings yet
- ATP Synthesis and Storage: Original ArticleDocument15 pagesATP Synthesis and Storage: Original ArticleRiley RilanNo ratings yet
- SHORT ESSAY - MetabolismDocument3 pagesSHORT ESSAY - MetabolismAllyzha AguilarNo ratings yet
- Lecture # 7 Pentose Phosphate PathwayDocument44 pagesLecture # 7 Pentose Phosphate PathwayDesy AnggrainiNo ratings yet
- The Pentose Phosphate Pathway in Health and Disease: Nature MetabolismDocument15 pagesThe Pentose Phosphate Pathway in Health and Disease: Nature Metabolismliliana-contrerasNo ratings yet
- Review Article: Thyrotoxic Periodic Paralysis: Clinical ChallengesDocument7 pagesReview Article: Thyrotoxic Periodic Paralysis: Clinical ChallengesRina ErlinaNo ratings yet
- Lintasan Pentosa FosfatDocument7 pagesLintasan Pentosa FosfatMUHAMMAD RIDONo ratings yet
- (CO8) Chemistry of VisionDocument7 pages(CO8) Chemistry of VisionCardoge DalisayNo ratings yet
- Disorders of Mitochondrial Long-Chain Fatty Acid oDocument15 pagesDisorders of Mitochondrial Long-Chain Fatty Acid o林怡伶No ratings yet
- Cold Spring Harb Symp Quant BiolDocument6 pagesCold Spring Harb Symp Quant BioleliNo ratings yet
- PHP2018Document25 pagesPHP2018radu nicolaeNo ratings yet
- HanooDocument8 pagesHanoomr samoNo ratings yet
- Pentose Phosphate PathwayDocument35 pagesPentose Phosphate PathwayRacheal MphandeNo ratings yet
- Artical Vit b6Document18 pagesArtical Vit b6sajjad khanNo ratings yet
- მიტოქონდრიის ფუნქცია და კიბოDocument1 pageმიტოქონდრიის ფუნქცია და კიბოEMD GROUPNo ratings yet
- Pyruvate Dehydrogenase LectureDocument34 pagesPyruvate Dehydrogenase LectureFerasKarajehNo ratings yet
- MetabolismDocument11 pagesMetabolismMituSamadderNo ratings yet
- Anaerobic Metabolism of Carbohydrates in RBC - Lectue XXIDocument4 pagesAnaerobic Metabolism of Carbohydrates in RBC - Lectue XXISaulNo ratings yet
- Glycolytic Pathways and Red Cell MetabolismDocument6 pagesGlycolytic Pathways and Red Cell Metabolismcaroline vaughanNo ratings yet
- Pterostilbene Reverses Palmitic Acid Mediated Insulin Resistance in HepG2 Cells by Reducing Oxidative Stress and Triglyceride AccumulationDocument14 pagesPterostilbene Reverses Palmitic Acid Mediated Insulin Resistance in HepG2 Cells by Reducing Oxidative Stress and Triglyceride Accumulationsajad ahmad MalikNo ratings yet
- Redox Biology: Research PaperDocument14 pagesRedox Biology: Research PaperSofiaNo ratings yet
- Ifosfamide PathwayDocument6 pagesIfosfamide PathwayDiana AhmadNo ratings yet
- PHD Thesis Oxidative StressDocument8 pagesPHD Thesis Oxidative Stressdawnrodriguezsaintpaul100% (1)
- Pharmacology of AutacoidsDocument13 pagesPharmacology of AutacoidsInocenteNo ratings yet
- Pentose Phosphate PathwayDocument32 pagesPentose Phosphate PathwayAbdimalik AliNo ratings yet
- Glucose As A Major Antioxidant - When, What For and Why It FailsDocument20 pagesGlucose As A Major Antioxidant - When, What For and Why It FailsDaniel GilbertoNo ratings yet
- Biochemistry CasesTeenage Weakling 2015Document5 pagesBiochemistry CasesTeenage Weakling 2015Neel KotrappaNo ratings yet
- 2010 11 E184-E193 Maria Delivoria-Papadopoulos and Peter J. MarroDocument12 pages2010 11 E184-E193 Maria Delivoria-Papadopoulos and Peter J. MarroAle JimenezNo ratings yet
- Molecules 27 07052Document20 pagesMolecules 27 07052Chris SmithNo ratings yet
- Note BiotechnologyDocument6 pagesNote BiotechnologyHala AlzuhairiNo ratings yet
- Carbohydrates Metabolism NotesDocument15 pagesCarbohydrates Metabolism NotesShaheenNo ratings yet
- Bba - Molecular Basis of Disease: SciencedirectDocument12 pagesBba - Molecular Basis of Disease: SciencedirectBiancaNo ratings yet
- Nutritional Support in Intensive Care Unit (ICU) PatientsDocument20 pagesNutritional Support in Intensive Care Unit (ICU) PatientssarahNo ratings yet
- Molecular Mechanism of Phenylhydrazine Induced Haematotoxicity: A ReviewDocument5 pagesMolecular Mechanism of Phenylhydrazine Induced Haematotoxicity: A ReviewJean FlorencondiaNo ratings yet
- Erythrocyte G6PD Activity and GSH Level As Risk Factors For Vascular Complications Among Type 2 Diabetics in Osogbo NigeriaDocument6 pagesErythrocyte G6PD Activity and GSH Level As Risk Factors For Vascular Complications Among Type 2 Diabetics in Osogbo NigeriaAudrey LyliNo ratings yet
- Oxygen Sensing, Homeostasis, and Disease: Review ArticleDocument11 pagesOxygen Sensing, Homeostasis, and Disease: Review ArticleMeidikaWulandariNo ratings yet
- Article Wjpps 1435648856Document11 pagesArticle Wjpps 1435648856Rio ImbaoNo ratings yet
- Surgical Treatment For Diffuse Coronary Artery DiseasesDocument17 pagesSurgical Treatment For Diffuse Coronary Artery DiseasesRedha FaridNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument8 pagesBiochemical and Biophysical Research CommunicationsAnnisa khairaNo ratings yet
- Lecture 4 GPCR-AA MetabolitesDocument22 pagesLecture 4 GPCR-AA MetabolitesDr. Sagheer AhmedNo ratings yet
- Final Exam Study ProblemsDocument6 pagesFinal Exam Study Problemsaboodh123No ratings yet
- Age-Dependent Levels of Protein Kinase Cs in Brain: Reduction of Endogenous Mechanisms of NeuroprotectionDocument18 pagesAge-Dependent Levels of Protein Kinase Cs in Brain: Reduction of Endogenous Mechanisms of NeuroprotectionAnonymous omr9p6uNo ratings yet
- Oxidative Stress in Cancer: ReviewDocument31 pagesOxidative Stress in Cancer: ReviewGabriela Alejandra Reina LaraNo ratings yet
- Cardiac GlycosidesDocument10 pagesCardiac GlycosidesGlupiaSprawaNo ratings yet
- Metabolic Changes Associated With Tumor Metastasis, Part 2: Mitochondria, Lipid and Amino Acid MetabolismDocument17 pagesMetabolic Changes Associated With Tumor Metastasis, Part 2: Mitochondria, Lipid and Amino Acid MetabolismUlfahRamadhaniNo ratings yet
- Original Contribution: Oxidative Stress in A Phenylketonuria Animal ModelDocument6 pagesOriginal Contribution: Oxidative Stress in A Phenylketonuria Animal ModelAndreea StefanNo ratings yet
- Biochemistry UW NotesDocument38 pagesBiochemistry UW NotesAnonymous lLiWgjFiNo ratings yet
- Lec.6 PPP Pathway PDFDocument5 pagesLec.6 PPP Pathway PDFhawraaNo ratings yet
- l10 Structure, Classification Biosynthesis of ProstaglandinsDocument22 pagesl10 Structure, Classification Biosynthesis of Prostaglandinsbilawal khanNo ratings yet
- (3.19) CarbosMetab (PPP, Other Sugars & GAGs)Document74 pages(3.19) CarbosMetab (PPP, Other Sugars & GAGs)pamelaNo ratings yet
- Genetically Determined Metabolism of Nicotine and Its Clinical SignificanceDocument7 pagesGenetically Determined Metabolism of Nicotine and Its Clinical Significanceerich70No ratings yet
- Xiong2011 2Document6 pagesXiong2011 2Alejandro SalazarNo ratings yet
- Final Warning A History of The New World Order by David Allen Rivera - SUMMARY / EXCERPTSDocument7 pagesFinal Warning A History of The New World Order by David Allen Rivera - SUMMARY / EXCERPTSKeith KnightNo ratings yet
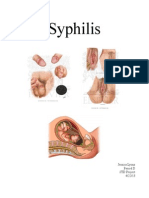
- Syphilis: Jessica Lyons Period D STD Project 4/22/13Document3 pagesSyphilis: Jessica Lyons Period D STD Project 4/22/13jesssielynnn33No ratings yet
- Tips No Naturally Cure Hemorrhoids FastDocument26 pagesTips No Naturally Cure Hemorrhoids FastK.l. DhanaNo ratings yet
- Wild and Ancient Fruit - Is It Really Small, Bitter, and Low in Sugar - Raw Food SOSDocument73 pagesWild and Ancient Fruit - Is It Really Small, Bitter, and Low in Sugar - Raw Food SOSVictor ResendizNo ratings yet
- Icd 10Document54 pagesIcd 10gexchaNo ratings yet
- Ultimate Guide To Surviving in The WildDocument175 pagesUltimate Guide To Surviving in The WildSal Ot100% (1)
- FM - Hse.020 Statistik HSE TahunanDocument2 pagesFM - Hse.020 Statistik HSE TahunanEka Nanda HermarianyNo ratings yet
- DEVPSY Reviewer - Chapters 1-6 PDFDocument19 pagesDEVPSY Reviewer - Chapters 1-6 PDFKathleen Anica SahagunNo ratings yet
- September 26, 2018 by Sagar Aryal: Antigen-Properties, Types and Determinants of AntigenicityDocument23 pagesSeptember 26, 2018 by Sagar Aryal: Antigen-Properties, Types and Determinants of AntigenicityEzekiel GantiedNo ratings yet
- 681 FullDocument6 pages681 FullKurnia AnharNo ratings yet
- ABC of Sexual Health: Taking A Sexual HistoryDocument10 pagesABC of Sexual Health: Taking A Sexual HistoryCristina PerezNo ratings yet
- Knowledge Regarding Immunization Among Mothers of Under Five ChildrenDocument3 pagesKnowledge Regarding Immunization Among Mothers of Under Five ChildrenEditor IJTSRDNo ratings yet
- Food Crisis in The World !Document6 pagesFood Crisis in The World !ChrissBright E. HermanNo ratings yet
- 24 VASQ Method VitA Intake 2006Document60 pages24 VASQ Method VitA Intake 2006aruniakpNo ratings yet
- Test Name Result Unit Reference Interval EndocrinolgyDocument1 pageTest Name Result Unit Reference Interval Endocrinolgygeetha kodaliNo ratings yet
- Postherpetic Neuralgia NerissaDocument18 pagesPostherpetic Neuralgia Nerissanerissa rahadianthiNo ratings yet
- Brief Notes Parish of St. RafaelDocument3 pagesBrief Notes Parish of St. RafaelAl F. Dela CruzNo ratings yet
- Presentation List Language Disorders in ChildrenDocument3 pagesPresentation List Language Disorders in Childrenramandeep kaurNo ratings yet
- Formative Gen EdDocument119 pagesFormative Gen EdDan GertezNo ratings yet
- ANNEX I ASEAN GP For The Negative List Ver 3.0 (14nov14)Document27 pagesANNEX I ASEAN GP For The Negative List Ver 3.0 (14nov14)blackcholoNo ratings yet
- Foot and Ankle Injuries Kylee Phillips - 0Document74 pagesFoot and Ankle Injuries Kylee Phillips - 0rizwan.mughal1997No ratings yet
- Probiotics, Prebiotics and The Gut Microbiota: Ilsi Europe Concise Monograph SeriesDocument40 pagesProbiotics, Prebiotics and The Gut Microbiota: Ilsi Europe Concise Monograph Seriesjimmy7forever100% (1)
- Patients' Perceptions of Barriers To Self-Managing Bipolar DisorderDocument10 pagesPatients' Perceptions of Barriers To Self-Managing Bipolar DisorderMilton MurilloNo ratings yet
- Demography Using Cemetery DataDocument5 pagesDemography Using Cemetery DatamjbdobleuNo ratings yet
- Clinical Schedule For Australian Dental Council Part 2 Exam CoachingDocument6 pagesClinical Schedule For Australian Dental Council Part 2 Exam CoachingdrrushikNo ratings yet
- Alcohol Research Paper OutlineDocument6 pagesAlcohol Research Paper Outlineafmctmvem100% (1)
- Apollo Excellence Report 2019 e VersionDocument289 pagesApollo Excellence Report 2019 e VersionrajNo ratings yet
- Gingival Inflammation in Mouth Breathing PatientsDocument5 pagesGingival Inflammation in Mouth Breathing PatientsCamiLunaNo ratings yet
- Phytochemical Analysis, Antimicrobial and Antioxidant Activities of Euphorbia Juss.) : An Unexplored Medicinal Herb Reported From CameroonDocument15 pagesPhytochemical Analysis, Antimicrobial and Antioxidant Activities of Euphorbia Juss.) : An Unexplored Medicinal Herb Reported From CameroonClara JamirNo ratings yet
- Hse Statistics Report Pp701 Hse f04 Rev.bDocument1 pageHse Statistics Report Pp701 Hse f04 Rev.bMohamed Mouner100% (1)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeFrom EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeRating: 4 out of 5 stars4/5 (1)
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeFrom EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeRating: 5 out of 5 stars5/5 (4)
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsFrom EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsRating: 4 out of 5 stars4/5 (146)
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsFrom EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsRating: 5 out of 5 stars5/5 (3)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeFrom EverandChemistry for Breakfast: The Amazing Science of Everyday LifeRating: 4.5 out of 5 stars4.5/5 (90)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolFrom EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolNo ratings yet
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableFrom EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableRating: 3.5 out of 5 stars3.5/5 (22)
- Meltdown: Nuclear disaster and the human cost of going criticalFrom EverandMeltdown: Nuclear disaster and the human cost of going criticalRating: 5 out of 5 stars5/5 (5)
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilFrom EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilRating: 5 out of 5 stars5/5 (1)
- Chemistry: a QuickStudy Laminated Reference GuideFrom EverandChemistry: a QuickStudy Laminated Reference GuideRating: 5 out of 5 stars5/5 (1)
- The Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookFrom EverandThe Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookNo ratings yet
- Handbook of Formulating Dermal Applications: A Definitive Practical GuideFrom EverandHandbook of Formulating Dermal Applications: A Definitive Practical GuideNo ratings yet
- Taste: Surprising Stories and Science About Why Food Tastes GoodFrom EverandTaste: Surprising Stories and Science About Why Food Tastes GoodRating: 3 out of 5 stars3/5 (20)
- Bioplastics: A Home Inventors HandbookFrom EverandBioplastics: A Home Inventors HandbookRating: 4 out of 5 stars4/5 (2)
- A Perfect Red: Empire, Espionage, and the Quest for the Color of DesireFrom EverandA Perfect Red: Empire, Espionage, and the Quest for the Color of DesireRating: 4 out of 5 stars4/5 (129)
- Formulating, Packaging, and Marketing of Natural Cosmetic ProductsFrom EverandFormulating, Packaging, and Marketing of Natural Cosmetic ProductsNo ratings yet
- Tribology: Friction and Wear of Engineering MaterialsFrom EverandTribology: Friction and Wear of Engineering MaterialsRating: 5 out of 5 stars5/5 (1)
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeFrom EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeRating: 5 out of 5 stars5/5 (1)
- The Periodic Table of Elements - Alkali Metals, Alkaline Earth Metals and Transition Metals | Children's Chemistry BookFrom EverandThe Periodic Table of Elements - Alkali Metals, Alkaline Earth Metals and Transition Metals | Children's Chemistry BookNo ratings yet