You might also like
- Screening For Cervical Cancer - UpToDateDocument40 pagesScreening For Cervical Cancer - UpToDateEvelin AlmeidaNo ratings yet
- OGTTDocument93 pagesOGTTapi-3769252No ratings yet
- Depression PowerpointDocument25 pagesDepression PowerpointLiezl Salatan AbuzoNo ratings yet
- 1 Glucose Responsive InsulinDocument2 pages1 Glucose Responsive InsulinJose J.No ratings yet
- Adaptasi Sel Beta Terhadap FruktosaDocument11 pagesAdaptasi Sel Beta Terhadap FruktosaEvan PermanaNo ratings yet
- 10 - 2020 - Vieira-Neto Et Al. Duration and Degree of Diet-Induced Metabolic Acidosis Prepartum Alters Tissue Responses To Insulin in Dairy CowsDocument20 pages10 - 2020 - Vieira-Neto Et Al. Duration and Degree of Diet-Induced Metabolic Acidosis Prepartum Alters Tissue Responses To Insulin in Dairy CowsRoney ZimpelNo ratings yet
- Physio 7Document12 pagesPhysio 7amjadNo ratings yet
- Journal of Steroid Biochemistry & Molecular Biology: A A A A A A A A A B C 1 A 1Document8 pagesJournal of Steroid Biochemistry & Molecular Biology: A A A A A A A A A B C 1 A 1Fuada Haerana RifaiNo ratings yet
- Aqp 7Document27 pagesAqp 7Blanca SolisNo ratings yet
- Hypergammaglobulinemia and Albumin Synthesis in The RabbitDocument2 pagesHypergammaglobulinemia and Albumin Synthesis in The RabbityanuararipratamaNo ratings yet
- Insulin InsightDocument8 pagesInsulin Insighttestreader1984No ratings yet
- Adaption of Sprague Dawley Rats To Long-Term Feeding of High Fat of High Fructose DietsDocument6 pagesAdaption of Sprague Dawley Rats To Long-Term Feeding of High Fat of High Fructose DietsmineortizvNo ratings yet
- InsulinDocument50 pagesInsulinTandin SonamNo ratings yet
- Asian Paci Fic Journal of Tropical MedicineDocument5 pagesAsian Paci Fic Journal of Tropical MedicineMuhammad Fuad MahfudNo ratings yet
- Improvement in Glycemia After Glucose or Insulin Overload in Leptin-Infused Rats Is Associated With Insulin-Related Activation of Hepatic Glucose MetabolismDocument6 pagesImprovement in Glycemia After Glucose or Insulin Overload in Leptin-Infused Rats Is Associated With Insulin-Related Activation of Hepatic Glucose MetabolismDaniel Gomez GalindoNo ratings yet
- Глюкагон мен глюкоза, инсулинDocument5 pagesГлюкагон мен глюкоза, инсулинArsen IzbanovNo ratings yet
- The Glucose Fatty-Acid Cycle. Its Role In... (Randle1963) Randle CycleDocument5 pagesThe Glucose Fatty-Acid Cycle. Its Role In... (Randle1963) Randle CycleFilip ZaleskiNo ratings yet
- Frank 2006Document8 pagesFrank 2006Rodrigo Martins RibeiroNo ratings yet
- Irregularity of Glycogen Synthase Homeostasis Complications Hepatic and Neuro DefectsDocument11 pagesIrregularity of Glycogen Synthase Homeostasis Complications Hepatic and Neuro DefectsEditor IJTSRDNo ratings yet
- Obesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierDocument9 pagesObesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. Fliersapphire_asaphNo ratings yet
- Insulin EndotextDocument37 pagesInsulin EndotextHendra HamzahNo ratings yet
- Insulin Under The Influence of Light: Ursino Gloria, Coppari RobertoDocument11 pagesInsulin Under The Influence of Light: Ursino Gloria, Coppari Robertoro111111No ratings yet
- Def Ronzo 2016Document32 pagesDef Ronzo 2016RachmatBimanjayaNo ratings yet
- GH Chap 78 - Insulin, Glucagon and DMDocument16 pagesGH Chap 78 - Insulin, Glucagon and DMShellz2428No ratings yet
- Nutrient Regulation of InsulinDocument16 pagesNutrient Regulation of InsulinNickolasMarllesNo ratings yet
- The Glucagon-Like Peptide-1 Receptor Agonist Oxyntomodulin Enhances Not Inhibit Gastric Emptying in MiceDocument9 pagesThe Glucagon-Like Peptide-1 Receptor Agonist Oxyntomodulin Enhances Not Inhibit Gastric Emptying in MiceFlavio SouzaNo ratings yet
- 7 Hormones of PancreasDocument29 pages7 Hormones of PancreasRana AbdullahNo ratings yet
- 2090 FullDocument9 pages2090 FullChloe WongNo ratings yet
- Fisiologi of PankreasDocument9 pagesFisiologi of PankreasSunardi BahriNo ratings yet
- Glucagon ReleaseDocument10 pagesGlucagon Releasesilvia pradoNo ratings yet
- Sem4a Insulin1995 JBCDocument4 pagesSem4a Insulin1995 JBCAntonella Farabelli SuarezNo ratings yet
- Beta Cell Specific Insr Deletion Promotes InsulinDocument22 pagesBeta Cell Specific Insr Deletion Promotes InsulinIda YulianaNo ratings yet
- DMJ 2021 0280Document23 pagesDMJ 2021 0280Helder CarvalhoNo ratings yet
- 1 Endocrinology of The Equine NeonateDocument10 pages1 Endocrinology of The Equine NeonateMariana CastañoNo ratings yet
- Grellina en DiabetesDocument19 pagesGrellina en DiabetesPaul SimononNo ratings yet
- Protein Kinase Cis Activated in From Rats: Glomeruli DiabeticDocument9 pagesProtein Kinase Cis Activated in From Rats: Glomeruli Diabeticapi-19757116No ratings yet
- Insulin 2 PDFDocument15 pagesInsulin 2 PDFAnonymous K9tmd7T0PzNo ratings yet
- Coffee and Caffeine Improve Insulin Sensitivity and Glucose Tolerance in C57BL 6J Mice Fed A High Fat DietDocument8 pagesCoffee and Caffeine Improve Insulin Sensitivity and Glucose Tolerance in C57BL 6J Mice Fed A High Fat DietIdaNo ratings yet
- Liaqat 2017Document8 pagesLiaqat 2017Hamza AldiriNo ratings yet
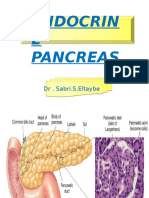
- Endocrin E Pancreas: DR - Sabri.S.EltaybeDocument43 pagesEndocrin E Pancreas: DR - Sabri.S.EltaybehamidNo ratings yet
- Green Tea Polyphenol Epigallocatechin-3-Gallate Ameliorates Insulin Resistance in Non-Alcoholic Fatty Liver Disease MiceDocument9 pagesGreen Tea Polyphenol Epigallocatechin-3-Gallate Ameliorates Insulin Resistance in Non-Alcoholic Fatty Liver Disease MiceFebyan AbotNo ratings yet
- Endocrinology 2003 p3958Document11 pagesEndocrinology 2003 p3958imperialscribdNo ratings yet
- Effects of Estrogen On Hyperglycemia and Liver DysDocument12 pagesEffects of Estrogen On Hyperglycemia and Liver DysdiskaNo ratings yet
- McGoldrick and Wheals 1989 SC GlucosamineDocument5 pagesMcGoldrick and Wheals 1989 SC GlucosamineAnaNo ratings yet
- 108038.1 20150304084446 CoveredDocument9 pages108038.1 20150304084446 Coveredعـَــٻاس مَـــشتاق عـَــليNo ratings yet
- Regulation of Insulin Receptor MetabolismDocument9 pagesRegulation of Insulin Receptor MetabolismShaheryar HasanNo ratings yet
- 100th Anniversary of The Discovery of Insulin Perspective Insulin and Adipose Tissue Fatty Acid MetabolismDocument18 pages100th Anniversary of The Discovery of Insulin Perspective Insulin and Adipose Tissue Fatty Acid MetabolismDamiano MolinaroNo ratings yet
- Obesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierDocument9 pagesObesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierernaataNo ratings yet
- Principles of Diabetes Mellitus - Ch3FinalDocument16 pagesPrinciples of Diabetes Mellitus - Ch3Finalphoenix sunNo ratings yet
- The Cellular Fate of Glucose and Its Relevance In-1-16Document16 pagesThe Cellular Fate of Glucose and Its Relevance In-1-16Yariela Amador MendietaNo ratings yet
- Direct Effect of Cholesterol On Insulin Secretion A Novel Mechanism For Pancreatic ?-Cell DysfunctionDocument11 pagesDirect Effect of Cholesterol On Insulin Secretion A Novel Mechanism For Pancreatic ?-Cell DysfunctionAsmat BurhanNo ratings yet
- CH 67Document14 pagesCH 67john mwangiNo ratings yet
- Physiology, Endocrine, Pancreas - StatPearls - NCBI BookshelfDocument5 pagesPhysiology, Endocrine, Pancreas - StatPearls - NCBI Bookshelfdrpaez27No ratings yet
- Glucose Transporters: Physiological and Pathological Roles: ReviewDocument5 pagesGlucose Transporters: Physiological and Pathological Roles: Review147 -Ashfia HashmiNo ratings yet
- Ghrelin Facilitates GLUT2-, SGLT1-and SGLT2-mediated Intestinal Glucose Transport in Goldfish (Carassius Auratus)Document16 pagesGhrelin Facilitates GLUT2-, SGLT1-and SGLT2-mediated Intestinal Glucose Transport in Goldfish (Carassius Auratus)Zulvina FaozanudinNo ratings yet
- Diabetes 2Document9 pagesDiabetes 2Adán LópezNo ratings yet
- GIP and GLP-1, The Two Incretin Hormones: Similarities and DifferencesDocument16 pagesGIP and GLP-1, The Two Incretin Hormones: Similarities and DifferencesYacine YacineNo ratings yet
- MBS2-K7-Pharmacology of Endocrine PancreasDocument55 pagesMBS2-K7-Pharmacology of Endocrine PancreasZoga Pratantia TohariNo ratings yet
- Efek Kandungan Serat Beras Analog Terhadap Ekspresi GLUT4 Otot Rangka Tikus DiabetesDocument8 pagesEfek Kandungan Serat Beras Analog Terhadap Ekspresi GLUT4 Otot Rangka Tikus DiabetesMelky oatsNo ratings yet
- Birkenfield 2014Document11 pagesBirkenfield 2014Bruno MoraesNo ratings yet
- Relationship To Muscle Triglyceride and w-3 Fatty Acids in Muscle PhospholipidDocument10 pagesRelationship To Muscle Triglyceride and w-3 Fatty Acids in Muscle PhospholipidAdeess AengNo ratings yet
- Leptin: Regulation and Clinical ApplicationsFrom EverandLeptin: Regulation and Clinical ApplicationsSam Dagogo-Jack, MDNo ratings yet
- A Comparative Study of Bhringarajadya Taila Nasya and Kumarika Varti Anjana in Timira W.S.R To Immature CataractDocument9 pagesA Comparative Study of Bhringarajadya Taila Nasya and Kumarika Varti Anjana in Timira W.S.R To Immature CataractAmit GargNo ratings yet
- Parasitology (Lect #2) TransDocument2 pagesParasitology (Lect #2) TransSherlyn Giban InditaNo ratings yet
- Chagaladya Ghrita A - Drug ReviewDocument8 pagesChagaladya Ghrita A - Drug ReviewEditor IJTSRDNo ratings yet
- Psychopathology/Problems in LivingDocument44 pagesPsychopathology/Problems in LivingDeadly ChillerNo ratings yet
- KernicterusDocument6 pagesKernicterusAirin Birthday GiftNo ratings yet
- Slaber Jaga Senin 3 Mei 2021Document7 pagesSlaber Jaga Senin 3 Mei 2021Louis MailuhuNo ratings yet
- Sandvikconstruction Dthkatalog LowresDocument44 pagesSandvikconstruction Dthkatalog Lowresroberto funezNo ratings yet
- Metabolic Syndrome Thesis PDFDocument5 pagesMetabolic Syndrome Thesis PDFafknawjof100% (2)
- Protocol ExampleDocument6 pagesProtocol ExampleWill RíosNo ratings yet
- CoronavirusDocument3 pagesCoronavirusĂhméđ ĂĺãshqãrNo ratings yet
- Pterygium Excision and Conjunctival AutograftDocument13 pagesPterygium Excision and Conjunctival Autograftgrace liwantoNo ratings yet
- Benefit of SportDocument6 pagesBenefit of SportNazmi JamilNo ratings yet
- Ebook Community Pharmacy Symptoms Diagnosis and Treatment PDF Full Chapter PDFDocument67 pagesEbook Community Pharmacy Symptoms Diagnosis and Treatment PDF Full Chapter PDFnancy.larsen721100% (26)
- Began in The 17th CenturyDocument82 pagesBegan in The 17th CenturyCharlene LunaNo ratings yet
- Tuberculosis NclexDocument3 pagesTuberculosis NclexMarinill SolimanNo ratings yet
- Acute Leukemia: Thirunavukkarasu MurugappanDocument22 pagesAcute Leukemia: Thirunavukkarasu MurugappanFelix Allen100% (1)
- Univesity of Sharjah 2023Document234 pagesUnivesity of Sharjah 2023Daysi Ramírez EscobarNo ratings yet
- TETANUSDocument41 pagesTETANUSruchikakaushal1910No ratings yet
- Surgery For Endocrinological Diseases and Malformations in ChildhoodDocument155 pagesSurgery For Endocrinological Diseases and Malformations in ChildhoodBeng DinNo ratings yet
- AnophthalmaDocument17 pagesAnophthalmaMarvellous OkonobohNo ratings yet
- The Long-Term Effects of Sexual Assault On Romantic RelationshipsDocument55 pagesThe Long-Term Effects of Sexual Assault On Romantic RelationshipsAndrés Yáñez ChavarriagaNo ratings yet
- Hypertension Concept MapDocument1 pageHypertension Concept Mapjyd parreñoNo ratings yet
- CDC Interim Reopening GuidanceDocument62 pagesCDC Interim Reopening GuidanceAlex GeliNo ratings yet
- Tirumala Tirupati Devasthanams (Official Booking Portal)Document1 pageTirumala Tirupati Devasthanams (Official Booking Portal)radhini anuNo ratings yet
- Radiation Proctitis Major Case Study Powerpoint FinalizedDocument22 pagesRadiation Proctitis Major Case Study Powerpoint Finalizedapi-634988720No ratings yet
- Why Do Adolescents DrinkDocument6 pagesWhy Do Adolescents DrinkchukisaliNo ratings yet
- NSTP-CWTS Syllabus Prelim-MidtermDocument16 pagesNSTP-CWTS Syllabus Prelim-MidtermKobe BryNo ratings yet
- PMUY (Introduction)Document3 pagesPMUY (Introduction)Lucifer RoxxNo ratings yet