You might also like
- MBB2 NeurologyDocument38 pagesMBB2 Neurologylizzy596No ratings yet
- Approach To Head CTDocument31 pagesApproach To Head CTElisabeth F. Ojha100% (2)
- Hiv Associated Hematological Malignasis - Marcus HentrichDocument239 pagesHiv Associated Hematological Malignasis - Marcus HentrichJaronjer1No ratings yet
- CSF and CNS InfectionsDocument9 pagesCSF and CNS InfectionsJorelyn Frias100% (1)
- Cranial NeuropathiesDocument74 pagesCranial NeuropathiesKevin Woodard100% (1)
- Muscle and Nervous Tissues LabDocument4 pagesMuscle and Nervous Tissues LabSip BioNo ratings yet
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesFrom EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNo ratings yet
- Small Animal Spinal Disorders Diagnosis and Surgery, 2nd Edition (Vetbooks - Ir)Document369 pagesSmall Animal Spinal Disorders Diagnosis and Surgery, 2nd Edition (Vetbooks - Ir)ghimboasaNo ratings yet
- (Biology) The LiverDocument7 pages(Biology) The LiverNiharikaNo ratings yet
- Spinal Cord Lesions: Jia Yan-Jie M.D PH.DDocument170 pagesSpinal Cord Lesions: Jia Yan-Jie M.D PH.Dapi-19916399No ratings yet
- LymphomaDocument78 pagesLymphomaastriwahyuni100% (4)
- Clinical Radio Biology - Previous Examination Papers Updated 040108Document12 pagesClinical Radio Biology - Previous Examination Papers Updated 040108sawiHijau100% (3)
- DISORDERS AND DISEASES CELL CYCLE - EllaDocument9 pagesDISORDERS AND DISEASES CELL CYCLE - EllaKennedy Fieldad Vagay100% (1)
- Lesson Plan Lumbar PunctureDocument25 pagesLesson Plan Lumbar PunctureValarmathi50% (4)
- Neuro Anatomy & Pathophysiology Laboratory Manual - 2 Semester, S. Y 2019 - 2020Document6 pagesNeuro Anatomy & Pathophysiology Laboratory Manual - 2 Semester, S. Y 2019 - 2020cris baligodNo ratings yet
- NeuroiflammationDocument9 pagesNeuroiflammationGabrielaNo ratings yet
- Genexglob ManualDocument23 pagesGenexglob ManualSurendran KrishnanNo ratings yet
- Reviews: The Anatomical and Cellular Basis of Immune Surveillance in The Central Nervous SystemDocument13 pagesReviews: The Anatomical and Cellular Basis of Immune Surveillance in The Central Nervous SystemGrupo AtlasNo ratings yet
- 2016 Neural Crest CellsDocument20 pages2016 Neural Crest Cells박다빈No ratings yet
- 10.1007@s12565 012 0166 6Document13 pages10.1007@s12565 012 0166 6San Phạm ĐìnhNo ratings yet
- Pathogenesis of The Central Nervous System InfectionDocument44 pagesPathogenesis of The Central Nervous System InfectionMira ApriliaNo ratings yet
- Introduction Into The Nervous System 2017-18Document58 pagesIntroduction Into The Nervous System 2017-18maodNo ratings yet
- The Cholinergic Anti-Inflammatory Pathway: A Missing Link in NeuroimmunomodulationDocument10 pagesThe Cholinergic Anti-Inflammatory Pathway: A Missing Link in NeuroimmunomodulationVandy IkraNo ratings yet
- Histo NervousDocument20 pagesHisto NervousALESSANDRA CELINo ratings yet
- 07 GeneralDocument13 pages07 GeneralMalinda KarunaratneNo ratings yet
- CNS - Infections F2022 OVER VIEWDocument18 pagesCNS - Infections F2022 OVER VIEWMadison MillwoodNo ratings yet
- Lecture 2nd Year NeuropathIntro - v3Document61 pagesLecture 2nd Year NeuropathIntro - v3asfsd asdsfasNo ratings yet
- January 1994, Volume 34, Number 1: Neurosurgery 1992-98 163 Intracranial Suprasellar AngiolipomaDocument8 pagesJanuary 1994, Volume 34, Number 1: Neurosurgery 1992-98 163 Intracranial Suprasellar AngiolipomaZdravko HeinrichNo ratings yet
- BME365S 25 Immune Overview 2020 HODocument49 pagesBME365S 25 Immune Overview 2020 HOBad BunnyNo ratings yet
- Blood Flow Regulation of Venous SystemDocument39 pagesBlood Flow Regulation of Venous SystemAmanuel MaruNo ratings yet
- Description of The Disease MeningitisDocument3 pagesDescription of The Disease MeningitisLance SilvaNo ratings yet
- L2 SBM Neuro - 2Document43 pagesL2 SBM Neuro - 2Sohil PatelNo ratings yet
- Common Neurologic Disorders: Krista M. GarnerDocument12 pagesCommon Neurologic Disorders: Krista M. GarnerTeda G.No ratings yet
- DC 1 PDFDocument25 pagesDC 1 PDFRebecca SandoicaNo ratings yet
- BIOS5130 Week 10 Lecture Part 1 SlidesDocument37 pagesBIOS5130 Week 10 Lecture Part 1 SlidesOkikiola JohnsonNo ratings yet
- Neuro PatDocument58 pagesNeuro Patakanksha nagarNo ratings yet
- 273 Full PDFDocument4 pages273 Full PDFAngelica Maria Rueda SerranoNo ratings yet
- Adverse Neural Tension WorkbookDocument29 pagesAdverse Neural Tension WorkbookCristi MansillaNo ratings yet
- Multiple SclerosisDocument3 pagesMultiple SclerosisVictorija Evania Lucille DeldioNo ratings yet
- Lecture Lesson 7. Cerebrospinal FluidDocument4 pagesLecture Lesson 7. Cerebrospinal FluidHANA LUNARIANo ratings yet
- Review Article: Neuroprotective Effects of Stem Cells in Ischemic StrokeDocument7 pagesReview Article: Neuroprotective Effects of Stem Cells in Ischemic StrokesalmaNo ratings yet
- NeurulationDocument7 pagesNeurulationFranTeodosioNo ratings yet
- Jessen2015 Article TheGlymphaticSystemABeginnerSGDocument17 pagesJessen2015 Article TheGlymphaticSystemABeginnerSGAhmad Al-HusainNo ratings yet
- I. II. Brain Stem III. Cerebellum IV. Diencephalon V. Cerebrum VI. Cranial NervesDocument60 pagesI. II. Brain Stem III. Cerebellum IV. Diencephalon V. Cerebrum VI. Cranial Nervesgaurav021No ratings yet
- ANAT CNS Support SystemsDocument52 pagesANAT CNS Support SystemsK SNo ratings yet
- Development of The Nervous System: Matt Velkey 454A Davison, Green Zone, Duke SouthDocument45 pagesDevelopment of The Nervous System: Matt Velkey 454A Davison, Green Zone, Duke SouthAbdimalik AliNo ratings yet
- NeurologyDocument19 pagesNeurologymerosaber622001No ratings yet
- 1 Nervous System OutlinesDocument45 pages1 Nervous System OutlinesTestingAccNo ratings yet
- 16 Pulp Lecture Varga EnglishDocument55 pages16 Pulp Lecture Varga EnglishRohmat SupriyadiNo ratings yet
- 4462 FullDocument10 pages4462 FullDokdem AjaNo ratings yet
- Research Open Access: Charlotte Schmitt, Nathalie Strazielle and Jean-François Ghersi-EgeaDocument15 pagesResearch Open Access: Charlotte Schmitt, Nathalie Strazielle and Jean-François Ghersi-Egeamihail.boldeanuNo ratings yet
- Neural CrestDocument33 pagesNeural CrestCarlo ManguilNo ratings yet
- Cerebral Edema Associated With MeningiomaDocument3 pagesCerebral Edema Associated With MeningiomaEstefania Ramirez CardonaNo ratings yet
- Blood Brain Barrier and Neurotransmitter 1Document43 pagesBlood Brain Barrier and Neurotransmitter 1Manirarora EtienneNo ratings yet
- Important QuestionsDocument38 pagesImportant Questionsas3941asNo ratings yet
- 2021 BBS2002 Case1 LectureDocument56 pages2021 BBS2002 Case1 LectureAnastasia Suzie AstrydeNo ratings yet
- Neurologic InvestigationsDocument31 pagesNeurologic InvestigationsISAH D. CONTORANo ratings yet
- 5 - Hydrocephalus 4 Course 2022Document44 pages5 - Hydrocephalus 4 Course 2022asaadwardNo ratings yet
- Lo SatuDocument5 pagesLo SatuRatihNo ratings yet
- Structure and Functions of Cells of The Nervous-1Document15 pagesStructure and Functions of Cells of The Nervous-1Attiya SohailNo ratings yet
- 200F-21 The Brain SVDocument20 pages200F-21 The Brain SVRachel SawhNo ratings yet
- 2017 - Dalkara - Cerebral MicrocirculationDocument5 pages2017 - Dalkara - Cerebral MicrocirculationmuelmiharNo ratings yet
- Infectious Diseases - Nervous System: Fcnlxa - St. Luke's College of NursingDocument5 pagesInfectious Diseases - Nervous System: Fcnlxa - St. Luke's College of NursingLAXA FRANCINENo ratings yet
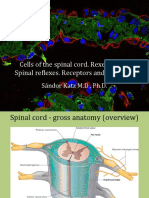
- Cells of The Spinal Cord. Rexed Zones. Spinal Reflexes. Receptors and EffectorsDocument34 pagesCells of The Spinal Cord. Rexed Zones. Spinal Reflexes. Receptors and Effectorsnnn nnnNo ratings yet
- Cellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8From EverandCellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8No ratings yet
- Green Life Cycle Science Butterfly Graphic OrganizerDocument1 pageGreen Life Cycle Science Butterfly Graphic OrganizerAlexia SalvadorNo ratings yet
- Anatomy& Physiology BY Adeniyi M. OlatoyeDocument59 pagesAnatomy& Physiology BY Adeniyi M. OlatoyeAdeniyi OlatoyeNo ratings yet
- Bio Assignment 1Document6 pagesBio Assignment 1ditucalan.ha2003No ratings yet
- PDFDocument4 pagesPDFacsNo ratings yet
- Hassan Et Al., 2024Document15 pagesHassan Et Al., 2024Herda CahyaningrumNo ratings yet
- Residu Kanamycin HPLCDocument2 pagesResidu Kanamycin HPLCAprelita Nurelli Dwiana100% (2)
- 155:411 Introduction To Biochemical Engineering Fall 2012 Exam 1 October 10, 2012 From 5:00-8:00 P.MDocument2 pages155:411 Introduction To Biochemical Engineering Fall 2012 Exam 1 October 10, 2012 From 5:00-8:00 P.Mmanthan2120% (1)
- Discovering PhotosynthesisDocument3 pagesDiscovering PhotosynthesisEarl CaridadNo ratings yet
- Science-8 - Q4 - WHLP 2021-2022Document8 pagesScience-8 - Q4 - WHLP 2021-2022Zerah ElsisuraNo ratings yet
- Physioex 1.1Document5 pagesPhysioex 1.1Tj MordeckiNo ratings yet
- Cmi CMBDocument7 pagesCmi CMBMary M. ToribioNo ratings yet
- PCR Troubleshooting-Part 1 "No Bands": Hints, Tips and Trouble Shooting For Molecular Biology TechniciansDocument6 pagesPCR Troubleshooting-Part 1 "No Bands": Hints, Tips and Trouble Shooting For Molecular Biology Technicianssam911101No ratings yet
- 71 Questions of Importance For Marine Conservation (ParsonsFavaro - Etal - 2014) PDFDocument10 pages71 Questions of Importance For Marine Conservation (ParsonsFavaro - Etal - 2014) PDFRoky Mountains HillNo ratings yet
- Yr 8 WK 2 Bio NoteDocument3 pagesYr 8 WK 2 Bio Notesedrick ocheNo ratings yet
- Conservation of BiodiversityDocument19 pagesConservation of BiodiversityAnkit AgarwalNo ratings yet
- Ex3 Abo Forward Grouping Group7Document19 pagesEx3 Abo Forward Grouping Group7marielle roseNo ratings yet
- 2 - Puberty-GirlsDocument40 pages2 - Puberty-Girlsapi-241585431No ratings yet
- The Sickle Cell Allele and Malaria IgcseDocument11 pagesThe Sickle Cell Allele and Malaria IgcseTanyaradzwa MugutaNo ratings yet
- Chapter 1.3 Structures and Functions of OrganellesDocument35 pagesChapter 1.3 Structures and Functions of OrganellesFairview PenangNo ratings yet
- Ishita MalikDocument10 pagesIshita MalikIshita malikNo ratings yet
- Bioprocess Technology: Industrial Importance of MicroorganismDocument4 pagesBioprocess Technology: Industrial Importance of MicroorganismAjay MahaichNo ratings yet
- Science 3-9 DLP Second QuarterDocument12 pagesScience 3-9 DLP Second QuarterCozette AtendidoNo ratings yet
- Daftar Pustaka PembahasanDocument2 pagesDaftar Pustaka PembahasanMaryam Fathiya RinthaniNo ratings yet
- Obesity 1. Basic Science of ObesityDocument10 pagesObesity 1. Basic Science of ObesitygraceNo ratings yet