You might also like
- Science 2Document3 pagesScience 2api-211640838No ratings yet
- Martinat Bottet, 2010 (Ed)Document11 pagesMartinat Bottet, 2010 (Ed)Andres SuarezNo ratings yet
- SEASONAL EFFECTS ON ESTRUS AND OVULATIONDocument7 pagesSEASONAL EFFECTS ON ESTRUS AND OVULATIONvidalbaezaNo ratings yet
- Amit Paper PublishedDocument4 pagesAmit Paper PublishedgnpobsNo ratings yet
- Moreira 2000Document9 pagesMoreira 2000PERLA CAROLINA ESPINOZA BRUNETNo ratings yet
- Effect of Ovariectomy On The Levels of Plasma Sex Hormones in Albino Rats E. A. Alagwu., R. O. NneliDocument5 pagesEffect of Ovariectomy On The Levels of Plasma Sex Hormones in Albino Rats E. A. Alagwu., R. O. NneliCaroline AgathaNo ratings yet
- Suzuki 2002Document7 pagesSuzuki 2002EveAriNo ratings yet
- 1 s2.0 S0022030217305416 MainahaDocument10 pages1 s2.0 S0022030217305416 MainahaBagas KarangNo ratings yet
- Effect of Estrogen Receptor Activation on Gonadotroph Progesterone Receptor Expression and Function in Young and Middle-Aged RatsDocument11 pagesEffect of Estrogen Receptor Activation on Gonadotroph Progesterone Receptor Expression and Function in Young and Middle-Aged RatsRoberto OrellanaNo ratings yet
- Che Cura 2010Document8 pagesChe Cura 2010camila andrea rivera diazNo ratings yet
- Determination of Ovulation Time in Bitches Based On Teasing, Vaginal Cytology, and Elisa For ProgesteroneDocument9 pagesDetermination of Ovulation Time in Bitches Based On Teasing, Vaginal Cytology, and Elisa For ProgesteroneAna Luísa SampaioNo ratings yet
- Induced Lactation in Cows and Heifers by Short-Term Treatment With Steroid HormonesDocument15 pagesInduced Lactation in Cows and Heifers by Short-Term Treatment With Steroid HormonesArfan YousafNo ratings yet
- Selection of A Dominant Follicle and Suppression of Follicular Growth in HeifersDocument13 pagesSelection of A Dominant Follicle and Suppression of Follicular Growth in HeifersGilson Antonio PessoaNo ratings yet
- Evidence For A Role of KISS-1/GPR54 System in Decreased Luteinizing Hormone (LH) Secretion in Fasted Prepubertal EwesDocument7 pagesEvidence For A Role of KISS-1/GPR54 System in Decreased Luteinizing Hormone (LH) Secretion in Fasted Prepubertal EwesAndrea CelisNo ratings yet
- Effect of Gonadotropinon Reproductive Performance of RabbitDocument11 pagesEffect of Gonadotropinon Reproductive Performance of RabbitCabrasDeVillaraureNo ratings yet
- I J V S: Nternational Ournal of Eterinary CienceDocument3 pagesI J V S: Nternational Ournal of Eterinary CienceDina AmaliaNo ratings yet
- Steotonin and dopamine relation with testoDocument7 pagesSteotonin and dopamine relation with testoupgratesleet704No ratings yet
- Gorbikrushanth_2017Document5 pagesGorbikrushanth_2017MiguelNo ratings yet
- PIIS00150282990060Document4 pagesPIIS00150282990060lilou.magnaboscoNo ratings yet
- Abdel Raouf 2015 Effect of Long Term Testosterone PRDocument9 pagesAbdel Raouf 2015 Effect of Long Term Testosterone PRangel chavezNo ratings yet
- Norstedt 1984Document6 pagesNorstedt 1984Malal QuechuaNo ratings yet
- tmp7495 TMPDocument6 pagestmp7495 TMPFrontiersNo ratings yet
- Adrenocortical Influences On Free Operant Avoidance BehaviorDocument9 pagesAdrenocortical Influences On Free Operant Avoidance BehaviorVíctor FuentesNo ratings yet
- Preclinical Acute Toxicity Studies and Rodent-Based Dosimetry Estimates of The Novel Sigma-1 Receptor Radiotracer (F) FPSDocument13 pagesPreclinical Acute Toxicity Studies and Rodent-Based Dosimetry Estimates of The Novel Sigma-1 Receptor Radiotracer (F) FPSirfanjadoonNo ratings yet
- 543 FullDocument6 pages543 FullantoniorNo ratings yet
- singh2001Document5 pagessingh2001f7jtbtdqnyNo ratings yet
- Herzog 2010Document7 pagesHerzog 2010Enzo German ZampiniNo ratings yet
- Psychoneuroendocrinology.: Et Al., Et Al., Et Al., Et Al., Et Al.Document11 pagesPsychoneuroendocrinology.: Et Al., Et Al., Et Al., Et Al., Et Al.MDNo ratings yet
- (6) O HCG é Mais Eficaz Do Que o Agonista GRH Buserelina Para Induzir a Primeira Ovulação Em Época de Reprodução Em ÉguasDocument6 pages(6) O HCG é Mais Eficaz Do Que o Agonista GRH Buserelina Para Induzir a Primeira Ovulação Em Época de Reprodução Em ÉguasIsabellaNo ratings yet
- Bartos Etal1998 SeasonalLevels IGF1CortisolReproductiveHormones MalePuduDocument6 pagesBartos Etal1998 SeasonalLevels IGF1CortisolReproductiveHormones MalePuduJosé Caro LagosNo ratings yet
- Telemetry: C-Fos and Dfosb Immunoreactivity in Rat Brain JT Cunningham Et AlDocument2 pagesTelemetry: C-Fos and Dfosb Immunoreactivity in Rat Brain JT Cunningham Et AlMicheal ChungNo ratings yet
- Fertility of Sows Injected With Exogenous Estradiol And-Or Gonadotropins To Control Post-Weaning Estrus Tcm96-154743Document6 pagesFertility of Sows Injected With Exogenous Estradiol And-Or Gonadotropins To Control Post-Weaning Estrus Tcm96-154743Bel Minda CostaNo ratings yet
- Ovarian Follicle Dynamics after Synchronization and SuperovulationDocument6 pagesOvarian Follicle Dynamics after Synchronization and SuperovulationAwang FaisalNo ratings yet
- Wildlife Research HorsesDocument7 pagesWildlife Research HorsesløzanNo ratings yet
- OlfatiDocument6 pagesOlfatiImran KhanNo ratings yet
- 1 s2.0 S1978301916303849 MainDocument4 pages1 s2.0 S1978301916303849 MainAnisaNo ratings yet
- Volume - 41 - 4 - 97 - 3 - MacacaDocument16 pagesVolume - 41 - 4 - 97 - 3 - MacacaAnindyaMustikaNo ratings yet
- Ijms 19 03457Document6 pagesIjms 19 03457efendirahimNo ratings yet
- Theriogenology: M.A. Driancourt, P. Cox, S. Rubion, G. Harnois-Milon, B. Kemp, N.M. SoedeDocument9 pagesTheriogenology: M.A. Driancourt, P. Cox, S. Rubion, G. Harnois-Milon, B. Kemp, N.M. SoedeAndres SuarezNo ratings yet
- Zinc Transport in Rabbit Tissues: Stems To at To To To Compartments Turnover RatesDocument8 pagesZinc Transport in Rabbit Tissues: Stems To at To To To Compartments Turnover RatesdugouskoNo ratings yet
- Lordosis Behavior in Male, Female and Androgenized Female RatsDocument13 pagesLordosis Behavior in Male, Female and Androgenized Female RatsHümay ÜnalNo ratings yet
- Acharjee et al. - 2017 - Ovaprim, a commercial spawning inducer, stimulatesDocument8 pagesAcharjee et al. - 2017 - Ovaprim, a commercial spawning inducer, stimulatesarup.acharjeeNo ratings yet
- Jvms 77 549Document8 pagesJvms 77 549ricepaddy1867No ratings yet
- Estron SulfatDocument7 pagesEstron SulfatMiswar KospalaNo ratings yet
- Epithelial Cycle: HybridDocument6 pagesEpithelial Cycle: HybridKamran JoyoNo ratings yet
- Jas 28 3 JAN0280030369Document6 pagesJas 28 3 JAN0280030369Roxana MariaNo ratings yet
- T.G.Cooper, C.Keck, U.Oberdieck and E.Nieschlag: Human Reproduction Vol.8 No.8 Pp. 1251-1258, 1993Document8 pagesT.G.Cooper, C.Keck, U.Oberdieck and E.Nieschlag: Human Reproduction Vol.8 No.8 Pp. 1251-1258, 1993Reski AdeliaNo ratings yet
- Weil Et Al Bio Letters 2006Document4 pagesWeil Et Al Bio Letters 2006zacharymweilNo ratings yet
- Scott E. Lukas Et Al - Phencyclidine-Analogue Self-Injection by The BaboonDocument5 pagesScott E. Lukas Et Al - Phencyclidine-Analogue Self-Injection by The BaboonBic0000No ratings yet
- Postoperative Pain Indicators in HorsesDocument13 pagesPostoperative Pain Indicators in HorsesAristoteles Esteban Cine VelazquezNo ratings yet
- Effect of Chronic High-Dose Exogenous Cortisol On Hippocampal Neuronal Number in Aged Nonhuman PrimatesDocument6 pagesEffect of Chronic High-Dose Exogenous Cortisol On Hippocampal Neuronal Number in Aged Nonhuman PrimatesFrontiersNo ratings yet
- Animreprod 9 1 52 PDFDocument9 pagesAnimreprod 9 1 52 PDFWAQAS SHAHIDNo ratings yet
- Hall 1954Document2 pagesHall 1954yom.moguelNo ratings yet
- Effect of Repeated Dual Superovulation Using FSH and PMSG+FSH On Ovulatory Response in Murrah BuffaloDocument8 pagesEffect of Repeated Dual Superovulation Using FSH and PMSG+FSH On Ovulatory Response in Murrah BuffaloGustav MolMedNo ratings yet
- Fasting Enhances Growth Hormone Secretion and Amplifies The Complex Rhythms of Growth Hormone Secretion in ManDocument9 pagesFasting Enhances Growth Hormone Secretion and Amplifies The Complex Rhythms of Growth Hormone Secretion in ManIvette Giselle LondoñoNo ratings yet
- 2009-79-1-3 - Aparitie Estrus - Lot1Document12 pages2009-79-1-3 - Aparitie Estrus - Lot1Anamaria Blaga PetreanNo ratings yet
- Flow CytometryDocument10 pagesFlow CytometryDaniel BarrigaNo ratings yet
- 1 s2.0 S0022030270864633 MainDocument12 pages1 s2.0 S0022030270864633 MainGaluh SuciningtyasNo ratings yet
- Reproduction in Domestic AnimalsFrom EverandReproduction in Domestic AnimalsH. H. ColeNo ratings yet
- Cost Per Test Listing of Immunoassay KitsDocument2 pagesCost Per Test Listing of Immunoassay Kitscocacola_thandaNo ratings yet
- Oral Contraceptive Reference ChartDocument6 pagesOral Contraceptive Reference Chartpdoan85No ratings yet
- AndrogensDocument19 pagesAndrogensMirza Shaharyar BaigNo ratings yet
- IPF Sanction Registry 20220914Document3 pagesIPF Sanction Registry 20220914Ryan AuliaNo ratings yet
- Gangguan Siklus Menstruasi Akibat Aktivitas Fisik Dan KecemasanDocument12 pagesGangguan Siklus Menstruasi Akibat Aktivitas Fisik Dan KecemasanSri AstutiNo ratings yet
- Gonadal Hormones and InhibitorsDocument10 pagesGonadal Hormones and InhibitorsCarlos NiñoNo ratings yet
- Superovulasi PDFDocument68 pagesSuperovulasi PDFHayatul IlmiNo ratings yet
- Anabolic Steroids for Sale in EuropeDocument2 pagesAnabolic Steroids for Sale in EuropelapiamaNo ratings yet
- The Biochemistry of Hormones ClassificationDocument45 pagesThe Biochemistry of Hormones ClassificationBadini ChanalNo ratings yet
- Hubungan Stress Dengan Gangguan Siklus Menstruasi Pada Siswi Kelas Xii Sma Karya RutengDocument7 pagesHubungan Stress Dengan Gangguan Siklus Menstruasi Pada Siswi Kelas Xii Sma Karya Rutengsondang nathasyaNo ratings yet
- Gonadotropins in ARTDocument37 pagesGonadotropins in ARTVarun DhallNo ratings yet
- Thyroid ReportDocument3 pagesThyroid ReportArman KhanNo ratings yet
- Sex Hormone Synthesis, Regulation, and Function - McMaster Pathophysiology ReviewDocument7 pagesSex Hormone Synthesis, Regulation, and Function - McMaster Pathophysiology ReviewWahyu W'cas PamungkasNo ratings yet
- Ichroma™ Brochure EliDocument4 pagesIchroma™ Brochure EliMeditec ecNo ratings yet
- Gonadotropin Releasing HormoneDocument9 pagesGonadotropin Releasing HormoneNTA UGC-NETNo ratings yet
- Function of Each GearsDocument6 pagesFunction of Each GearsRacco Roy0% (1)
- Price List of Sensetech S300 Immunoflorescence Analyzer Uk: Valid Till Dec 31, 2021Document2 pagesPrice List of Sensetech S300 Immunoflorescence Analyzer Uk: Valid Till Dec 31, 2021muhammad israeel100% (1)
- Jawaban Lo Kelompok (JLK) Scenario 3 Kelompok Tutorial 5Document9 pagesJawaban Lo Kelompok (JLK) Scenario 3 Kelompok Tutorial 5NOVI UMAMI UMAMINo ratings yet
- LH 1st by Dr. Manisha Jain - Jindal IVF ChandigarhDocument37 pagesLH 1st by Dr. Manisha Jain - Jindal IVF ChandigarhJindal IVF ChandigarhNo ratings yet
- Hormonal Contraceptives in CanadaDocument3 pagesHormonal Contraceptives in CanadaNida Saghir AhmedNo ratings yet
- Post Cycle TherapyDocument2 pagesPost Cycle TherapyXBBXNo ratings yet
- Certificate of Analysis: Liaison Control Thyroid 3Document1 pageCertificate of Analysis: Liaison Control Thyroid 3Khaled AlkhawaldehNo ratings yet
- Hormone Therapy: The Basics: What Are Hormones and How Are They Used As Therapy?Document4 pagesHormone Therapy: The Basics: What Are Hormones and How Are They Used As Therapy?Ubaidillah Romadlon AlfairuziNo ratings yet
- Product Active IngredientDocument2 pagesProduct Active IngredientOuamrouche SouheilNo ratings yet
- Which OCP Is Best Handout PDFDocument4 pagesWhich OCP Is Best Handout PDFMarianna LeeNo ratings yet
- MR NITIN PDFDocument2 pagesMR NITIN PDFVenkat Nitin GuttaNo ratings yet
- THEORIES OF ANDROPAUSEDocument55 pagesTHEORIES OF ANDROPAUSESyam RJNo ratings yet
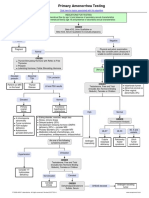
- Primary Amenorrhea Testing AlgorithmDocument1 pagePrimary Amenorrhea Testing AlgorithmfarmasiNo ratings yet
- Infographics of Endocrine SystemDocument2 pagesInfographics of Endocrine System2086151No ratings yet
- Testosterone Cypionate ProtocolDocument2 pagesTestosterone Cypionate ProtocolWidfdsafdsaNo ratings yet