You might also like
- Arboviruses in ChinaDocument21 pagesArboviruses in ChinaChris LicínioNo ratings yet
- UntitledDocument26 pagesUntitledLaura SanchezNo ratings yet
- Fisiopatología 1Document8 pagesFisiopatología 1CARLOS MONCAYO DIAZNo ratings yet
- Factors Affecting SARS CoV 2 (COVID-19) PandemicDocument32 pagesFactors Affecting SARS CoV 2 (COVID-19) PandemicTahir AliNo ratings yet
- Viruses 16 00214 v2Document16 pagesViruses 16 00214 v2valdirbrazsNo ratings yet
- Solignat Et Al. - 2009 - Replication Cycle of Chikungunya A Re-Emerging ArbovirusDocument15 pagesSolignat Et Al. - 2009 - Replication Cycle of Chikungunya A Re-Emerging ArbovirusLaise Eduarda Paixão de MoraesNo ratings yet
- Marine Natural Products in The Battle Against Dengue, ZikaDocument17 pagesMarine Natural Products in The Battle Against Dengue, ZikaBeatriz HenkelsNo ratings yet
- Zika Virus Transmission by Brazilian Aedes Temperature-DependentDocument14 pagesZika Virus Transmission by Brazilian Aedes Temperature-Dependentthais chouin-carneiroNo ratings yet
- Alphavirus Identification in Neotropical BatsDocument11 pagesAlphavirus Identification in Neotropical BatsItrux jonathan cisnerosNo ratings yet
- Oropouche Virus As An Emerging Cause of Acute Febrile Illness in ColombiaDocument13 pagesOropouche Virus As An Emerging Cause of Acute Febrile Illness in ColombiaVictor CostaNo ratings yet
- Maulidia Tugas B. InggrisDocument3 pagesMaulidia Tugas B. InggrisMauli LidyaNo ratings yet
- 2019 - 00 - Prudhomme & AlDocument12 pages2019 - 00 - Prudhomme & Aljcg91.973No ratings yet
- Contributions of Veterinary Profession To Management of COVID - 19 Pandemic - Epashupalan Apr 2021Document3 pagesContributions of Veterinary Profession To Management of COVID - 19 Pandemic - Epashupalan Apr 2021Deepak CNo ratings yet
- 2020 - Vector Competence of Aedes Aegypti, Aedes Albopictus and Culex Quinquefasciatus Mosquitoes For Mayaro VirusDocument13 pages2020 - Vector Competence of Aedes Aegypti, Aedes Albopictus and Culex Quinquefasciatus Mosquitoes For Mayaro VirusLúcia ReisNo ratings yet
- The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and ControlDocument29 pagesThe COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and ControlkusumawardatiNo ratings yet
- FULLTEXT01Document16 pagesFULLTEXT01docfallamine8678No ratings yet
- Zika Chikungunya and Dengue. The Causes and Threats of New and Reemerging Arboviral DiseasesDocument8 pagesZika Chikungunya and Dengue. The Causes and Threats of New and Reemerging Arboviral DiseasesEduarda QuartinNo ratings yet
- Petersen Human Monkeypox Rev ID Clin NA 2019Document17 pagesPetersen Human Monkeypox Rev ID Clin NA 2019thaistoledofinkNo ratings yet
- Corona VirusDocument21 pagesCorona VirusAnand JeeNo ratings yet
- 3luminex DengueDocument15 pages3luminex DengueMary Paula EstiveNo ratings yet
- Articulo VirologiaDocument12 pagesArticulo VirologiaValentina Maria Franco MartinezNo ratings yet
- Are Pangolins The Intermediate Host of The 2019 Novel Coronavirus (2019-Ncov) ?Document30 pagesAre Pangolins The Intermediate Host of The 2019 Novel Coronavirus (2019-Ncov) ?jm852kNo ratings yet
- vms3 434Document9 pagesvms3 434ali sherNo ratings yet
- Humanmonkeypox: Epidemiologic and Clinical Characteristics, Diagnosis, and PreventionDocument17 pagesHumanmonkeypox: Epidemiologic and Clinical Characteristics, Diagnosis, and PreventionAlejandro GraterolNo ratings yet
- 30 39Document10 pages30 39Helena Putri SiraitNo ratings yet
- A Review of The Monkeypox Virus and A Recent Outbreak of Skin Rash Disease in Nigeria: Kabuga and El ZowalatyDocument9 pagesA Review of The Monkeypox Virus and A Recent Outbreak of Skin Rash Disease in Nigeria: Kabuga and El ZowalatyNers LollipopNo ratings yet
- ZIKA VIRUS-An Update With Indian PerspectiveDocument5 pagesZIKA VIRUS-An Update With Indian PerspectiveAdvanced Research PublicationsNo ratings yet
- The Prevalence of Parasitic Protozoan Diseases in Iraq, 2016Document5 pagesThe Prevalence of Parasitic Protozoan Diseases in Iraq, 2016Jeremia AnkesaNo ratings yet
- Transmission Dynamics of Crimean-Congo HaemorrhagiDocument17 pagesTransmission Dynamics of Crimean-Congo Haemorrhagiali zohaibNo ratings yet
- Oropouche Virus As An Emerging Cause of Acute Febrile Illness in ColombiaDocument14 pagesOropouche Virus As An Emerging Cause of Acute Febrile Illness in Colombiajuliana22004No ratings yet
- Surveillance and Management of Chikungunya DiseaseDocument11 pagesSurveillance and Management of Chikungunya Diseasegiggs_libraNo ratings yet
- Mosquito As VectorDocument11 pagesMosquito As VectorSRI KATHIRNo ratings yet
- Seroprevalence of Cattle Respiratory Viral Pathongens in Paranatinga, Mato Grosso State, BrazilDocument5 pagesSeroprevalence of Cattle Respiratory Viral Pathongens in Paranatinga, Mato Grosso State, BrazilIJAERS JOURNALNo ratings yet
- Potential Zoonotic Bat-Borne Disease in Indonesia - Yan Newsletter Jan 2022Document12 pagesPotential Zoonotic Bat-Borne Disease in Indonesia - Yan Newsletter Jan 2022YanMardianNo ratings yet
- PIIS2405844021025998Document11 pagesPIIS2405844021025998Luthvia AnnisaNo ratings yet
- Souza-Neto Powell Bonizzoni. 2019. Aedes Aegypti Vector Competence Studies. A Review.Document19 pagesSouza-Neto Powell Bonizzoni. 2019. Aedes Aegypti Vector Competence Studies. A Review.Laura FloresNo ratings yet
- Non-Target Effects of Chemical Malaria Vector Control OnDocument12 pagesNon-Target Effects of Chemical Malaria Vector Control Ongerges8723No ratings yet
- Travel Medicine and Infectious Disease: ReviewDocument10 pagesTravel Medicine and Infectious Disease: Reviewimamsantos1191No ratings yet
- Ebola Virus Disease: A Manual for EVD ManagementFrom EverandEbola Virus Disease: A Manual for EVD ManagementMarta LadoNo ratings yet
- Clinical and Epidemiological Characteristics of Rotavirus and Norovirus Infection in Children in Andijan RegionDocument5 pagesClinical and Epidemiological Characteristics of Rotavirus and Norovirus Infection in Children in Andijan RegionCentral Asian StudiesNo ratings yet
- Human MonkeypoxDocument17 pagesHuman MonkeypoxJessica C. S.No ratings yet
- Zoonotic Diseases 1Document2 pagesZoonotic Diseases 1MelNo ratings yet
- Upwork 1Document118 pagesUpwork 1Yellow MellowNo ratings yet
- Fmicb 11 631736Document16 pagesFmicb 11 631736Herick Lyncon Antunes RodriguesNo ratings yet
- (Part 2) Potential Zoonotic Bat-Borne Disease in Indonesia - Yan Newsletter Feb 2022Document7 pages(Part 2) Potential Zoonotic Bat-Borne Disease in Indonesia - Yan Newsletter Feb 2022YanMardianNo ratings yet
- Jvi 01778-22Document14 pagesJvi 01778-22Paulo BorattoNo ratings yet
- A Review of The Monkeypox Virus and A Recent Outbreak of Skin Rash Disease in NigeriaDocument9 pagesA Review of The Monkeypox Virus and A Recent Outbreak of Skin Rash Disease in NigeriaNadia Desanti RachmatikaNo ratings yet
- Ji Et Al-2020-Journal of Medical Virology PDFDocument29 pagesJi Et Al-2020-Journal of Medical Virology PDFEwis Romero HillsNo ratings yet
- Virus Survival in The Environment: E.C. Pirtleandg.W. BeranDocument16 pagesVirus Survival in The Environment: E.C. Pirtleandg.W. BeranSoul LyricsNo ratings yet
- Dengue LancetDocument16 pagesDengue LancetJorge Ricardo Uchuya GómezNo ratings yet
- Incidence of Intestinal Helminthes Among HIV Patients at Kampala International University Teaching Hospital, Uganda.Document12 pagesIncidence of Intestinal Helminthes Among HIV Patients at Kampala International University Teaching Hospital, Uganda.KIU PUBLICATION AND EXTENSIONNo ratings yet
- Zoonotic and Reverse Zoonotic Events of SARS CoV 2 and Their Impact On Global HealthDocument15 pagesZoonotic and Reverse Zoonotic Events of SARS CoV 2 and Their Impact On Global HealthvivianNo ratings yet
- Transmisi Virus DengueDocument8 pagesTransmisi Virus DengueLiza Amanda SaphiraNo ratings yet
- 3.01.05 CCHFDocument8 pages3.01.05 CCHFkcstincasNo ratings yet
- Study of The Factors Associated With Domestic Ruminants Rift Valley Fever Epidemic in Mauritania, 2020Document6 pagesStudy of The Factors Associated With Domestic Ruminants Rift Valley Fever Epidemic in Mauritania, 2020International Journal of Innovative Science and Research TechnologyNo ratings yet
- Risk Factors For Chikungunya Outbreak in Kebridhar City, Somali Ethiopia, 2019. Unmatched Case-Control StudyDocument21 pagesRisk Factors For Chikungunya Outbreak in Kebridhar City, Somali Ethiopia, 2019. Unmatched Case-Control StudyNigatu AdmasuNo ratings yet
- Impact of Rotavirus Vaccine Introduction in Abidjan Côte D'ivoireDocument4 pagesImpact of Rotavirus Vaccine Introduction in Abidjan Côte D'ivoireArmel BheratNo ratings yet
- Review COVID-19 NEJMDocument13 pagesReview COVID-19 NEJMElielNo ratings yet
- Chikungunya Viral Infections An Emerging ProblemDocument6 pagesChikungunya Viral Infections An Emerging ProblemMike zombieNo ratings yet
- Approach To STDS2Document56 pagesApproach To STDS2Daniel ThomasNo ratings yet
- DotsDocument5 pagesDotsYusephAwangSanaaniNo ratings yet
- Doctor's Order & Lab ResultsDocument4 pagesDoctor's Order & Lab ResultsKaloy Kamao100% (1)
- Uiip Flu Fact SheetDocument4 pagesUiip Flu Fact Sheetapi-277123092No ratings yet
- Chapter 43: Nursing Care of A Family When A Child Has An Infectious DisorderDocument12 pagesChapter 43: Nursing Care of A Family When A Child Has An Infectious DisorderAlyssaGrandeMontimorNo ratings yet
- Dyshidrotic EczemaDocument2 pagesDyshidrotic EczemaatikaNo ratings yet
- Infection Control, Safety First Aid and Personal WellnessDocument9 pagesInfection Control, Safety First Aid and Personal WellnessJacinta MalamionNo ratings yet
- National Leprosy Eradication Programme (Nlep)Document34 pagesNational Leprosy Eradication Programme (Nlep)Balaji KrishnanNo ratings yet
- Am. J. Epidemiol.-1999-McMichael-887-97Document11 pagesAm. J. Epidemiol.-1999-McMichael-887-97juanpaNo ratings yet
- History of Biostatistics: Correspondence ToDocument5 pagesHistory of Biostatistics: Correspondence ToHenrique MartinsNo ratings yet
- Lifeline Quartely: IamiwillDocument12 pagesLifeline Quartely: IamiwillGrace TuvakasigaNo ratings yet
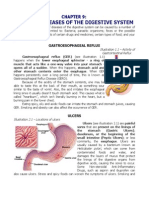
- Diseases of The Digestive SystemDocument5 pagesDiseases of The Digestive SystemG1N0G4M3No ratings yet
- Brochure DepressionDocument2 pagesBrochure Depressionel milanesasNo ratings yet
- Lippincott - S Illustrated Q - A Review of Microbiology - Immunology VirusDocument17 pagesLippincott - S Illustrated Q - A Review of Microbiology - Immunology VirusGoose ChapmanNo ratings yet
- Prevalensi Infeksi Cacing Usus Soil Transmitted Helminths Pada Orang Dewasa Di Sulawesi UtaraDocument4 pagesPrevalensi Infeksi Cacing Usus Soil Transmitted Helminths Pada Orang Dewasa Di Sulawesi UtaraSahrul hamidNo ratings yet
- Malaria 8Document35 pagesMalaria 8Fuad Nasir 0913No ratings yet
- Case Presentation Acute Tonsillopharyngitis: Pamantasan NG Cabuayo College of NursingDocument35 pagesCase Presentation Acute Tonsillopharyngitis: Pamantasan NG Cabuayo College of Nursingreymarhay50% (2)
- Epilepsia - UpToDateDocument28 pagesEpilepsia - UpToDateMiguelQuispeZeladaNo ratings yet
- 20-P2S2 Examination PapersDocument26 pages20-P2S2 Examination PapersSENo ratings yet
- Mu 089Document4 pagesMu 089Rahul RaiNo ratings yet
- Arleeluck C2395-Stu - Cancer Brochure TemplateDocument3 pagesArleeluck C2395-Stu - Cancer Brochure Templateapi-552283875No ratings yet
- NURSING CARE PLAN Impaired Breathing PatternDocument3 pagesNURSING CARE PLAN Impaired Breathing PatternChie Hyun-AeNo ratings yet
- Asepsis and Infection Control (Week 4)Document7 pagesAsepsis and Infection Control (Week 4)JEREMY MAKALINTALNo ratings yet
- Capstone Case F 4Document6 pagesCapstone Case F 4Ameer HamzaNo ratings yet
- Measles: The New Red Scare: by Oman YstrianykDocument20 pagesMeasles: The New Red Scare: by Oman YstrianykManuel AlbaNo ratings yet
- Epidemiological Traid JayaDocument25 pagesEpidemiological Traid JayajayalakshmiNo ratings yet
- Severe Acute Respiratory Syndrome (Sars) : Department of Microbiology 12 January 2015Document24 pagesSevere Acute Respiratory Syndrome (Sars) : Department of Microbiology 12 January 2015revathidadam55555No ratings yet
- Case Control StudyDocument34 pagesCase Control StudyRENJULAL100% (1)
- Diabetic RetinopathyDocument13 pagesDiabetic RetinopathyMuhammad Rhema0% (1)
- Traveller DiarheaDocument43 pagesTraveller Diarheatri farmaniNo ratings yet