You might also like
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967From EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterNo ratings yet
- TMP F204Document2 pagesTMP F204FrontiersNo ratings yet
- Baratta2002-Somatostatin LTP Dentate GyrusDocument9 pagesBaratta2002-Somatostatin LTP Dentate Gyrusiulia andreeaNo ratings yet
- RAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeDocument14 pagesRAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeFedrmNo ratings yet
- Lin Etal 2003Document9 pagesLin Etal 2003林瑜璿No ratings yet
- Cyclic AMP-mediated Inhibition of Noradrenaline-Induced Contraction Ca Influx in Guinea-Pig Vas DeferensDocument12 pagesCyclic AMP-mediated Inhibition of Noradrenaline-Induced Contraction Ca Influx in Guinea-Pig Vas DeferensssmorgNo ratings yet
- BF02462837Document4 pagesBF02462837ttqnhu.rhmNo ratings yet
- Wendy W. Wu, C. Savio Chan, D. James Surmeier and John F. DisterhoftDocument13 pagesWendy W. Wu, C. Savio Chan, D. James Surmeier and John F. DisterhoftFedrmNo ratings yet
- Muscarinic Receptors Modulate N-, P-, and L-Type Ca2+ Currents in Rat Striatal Neurons Through Parallel PathwaysDocument12 pagesMuscarinic Receptors Modulate N-, P-, and L-Type Ca2+ Currents in Rat Striatal Neurons Through Parallel PathwaysJunmajNo ratings yet
- Christian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionDocument13 pagesChristian Ducreux and Jean-Jacques Puizillout - A-Current Modifies The Spike of C-Type Neurones in The Rabbit Nodose GanglionFedrmNo ratings yet
- R.A. North and T. Tokimasa - Depression of Calcium-Dependent Potassium Conductance of Guinea-Pig Myenteric Neurones by Muscaric AgonistsDocument14 pagesR.A. North and T. Tokimasa - Depression of Calcium-Dependent Potassium Conductance of Guinea-Pig Myenteric Neurones by Muscaric AgonistsFedrmNo ratings yet
- Long-Term Potentiation of Intrinsic Excitability at The Mossy Fiber - Granule Cell Synapse of Rat CerebellumDocument9 pagesLong-Term Potentiation of Intrinsic Excitability at The Mossy Fiber - Granule Cell Synapse of Rat CerebellumJar JarNo ratings yet
- Soluble A Peptide Increases Excitatory Neurotransmission and Induces Epileptiform Activity in Hippocampal NeuronsDocument15 pagesSoluble A Peptide Increases Excitatory Neurotransmission and Induces Epileptiform Activity in Hippocampal NeuronsClaudia Alejandra Lopez ToroNo ratings yet
- Bear&KirkwoodDocument11 pagesBear&KirkwoodswagatarcNo ratings yet
- Effects of Low Concentrations of 4-Aminopyridine On CA1 Pyramidal Cells of The HippocampusDocument18 pagesEffects of Low Concentrations of 4-Aminopyridine On CA1 Pyramidal Cells of The HippocampusJortegloriaNo ratings yet
- T.G.J. Allen and G. Burnstock - Intracellular Studies of The Electrophysiological Properties of Cultured Intracardiac Neurones of The Guinea-PigDocument18 pagesT.G.J. Allen and G. Burnstock - Intracellular Studies of The Electrophysiological Properties of Cultured Intracardiac Neurones of The Guinea-PigLuammmNo ratings yet
- Delayed Increase of Acetylcholine Quantal Size Induced by THE Activity-Dependent Release of Endogenous CGRP But Not Atp in Neuromuscular JunctionsDocument25 pagesDelayed Increase of Acetylcholine Quantal Size Induced by THE Activity-Dependent Release of Endogenous CGRP But Not Atp in Neuromuscular JunctionsgarridotfNo ratings yet
- Biophysj00107 0239Document6 pagesBiophysj00107 0239aydinmemmedov9009No ratings yet
- Taniguchi 1983Document10 pagesTaniguchi 1983Giulia AndreeaNo ratings yet
- Membrane Stretch Affects Gating Modes of A Skeletal Muscle Sodium ChannelDocument17 pagesMembrane Stretch Affects Gating Modes of A Skeletal Muscle Sodium ChannelMichael MaierNo ratings yet
- Novel Strategy For Measuring Creatine Kinase Reaction Rate in The in Vivo HeartDocument10 pagesNovel Strategy For Measuring Creatine Kinase Reaction Rate in The in Vivo HeartYoung-Hoon SungNo ratings yet
- Lei 1998Document12 pagesLei 19980hitk0No ratings yet
- Burn Injury Decreases Myocardial Na-K-Atpase Activity: Role of PKC InhibitionDocument9 pagesBurn Injury Decreases Myocardial Na-K-Atpase Activity: Role of PKC InhibitionChristine Notoningtiyas SantosoNo ratings yet
- Rapid Temperature Jumps Using Infrared Diode Laser for Patch-Clamp StudiesDocument9 pagesRapid Temperature Jumps Using Infrared Diode Laser for Patch-Clamp StudiesAldo Alexander CamachoNo ratings yet
- TMP 75 AADocument9 pagesTMP 75 AAFrontiersNo ratings yet
- Jvms 83 997Document7 pagesJvms 83 997Blanca A SerranoNo ratings yet
- Takuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1Document18 pagesTakuya Yasui Et Al - Dynamic Synapses As Archives of Synaptic History: State-Dependent Redistribution of Synaptic Efficacy in The Rat Hippocampal CA1FedrmNo ratings yet
- Activation of Medial Prefrontal Cortex by Phencyclidine Is Mediated Via A Hippocampo-Prefrontal PathwayDocument7 pagesActivation of Medial Prefrontal Cortex by Phencyclidine Is Mediated Via A Hippocampo-Prefrontal PathwayCortate15gNo ratings yet
- European Journal of PharmacologyDocument6 pagesEuropean Journal of Pharmacologyliska ramdanawatiNo ratings yet
- Adenylyl Cyclase in The Heart: An Enzymocytochemical and Immunocytochemical ApproachDocument6 pagesAdenylyl Cyclase in The Heart: An Enzymocytochemical and Immunocytochemical ApproachJoNo ratings yet
- Supporting Information: Roy Et Al. 10.1073/pnas.1708261114Document5 pagesSupporting Information: Roy Et Al. 10.1073/pnas.1708261114Jeffer ZevallosNo ratings yet
- Nitric Oxide, CGMP, and Hormone Regulation of Active Sodium TransportDocument6 pagesNitric Oxide, CGMP, and Hormone Regulation of Active Sodium TransportMichael Daley100% (1)
- Kondo 2000, Putative Ryanodine Receptors in The Sarcolemma of Ventricular Myocytes.Document7 pagesKondo 2000, Putative Ryanodine Receptors in The Sarcolemma of Ventricular Myocytes.Alessio LissoniNo ratings yet
- Brozoski 2005Document6 pagesBrozoski 2005Caesar Catalin CaratasuNo ratings yet
- TMP C32Document11 pagesTMP C32FrontiersNo ratings yet
- TMP 9 DA2Document11 pagesTMP 9 DA2FrontiersNo ratings yet
- Supplement Material Methods Cardiomyocyte IsolationDocument12 pagesSupplement Material Methods Cardiomyocyte IsolationhNo ratings yet
- Cordycepin - CA1 Membrane HyperpolarizationDocument5 pagesCordycepin - CA1 Membrane Hyperpolarizationttqnhu.rhmNo ratings yet
- Exercise Induces Muscle Fiber Type SwitchingDocument10 pagesExercise Induces Muscle Fiber Type SwitchingbookNo ratings yet
- BHP 311Document9 pagesBHP 311antoniorNo ratings yet
- A. Reyes Et Al - Passive Properties of Neostriatal Neurons During Potassium Conductance BlockadeDocument15 pagesA. Reyes Et Al - Passive Properties of Neostriatal Neurons During Potassium Conductance BlockadeFedrmNo ratings yet
- Plasticity in The Human Central Nervous SystemDocument15 pagesPlasticity in The Human Central Nervous SystemMarcelo LugonNo ratings yet
- Letter To The Editor: Doi: 10.3967/bes2017.125Document5 pagesLetter To The Editor: Doi: 10.3967/bes2017.125Ion CorbuNo ratings yet
- Control of Airway Smooth Muscle Tone. I - Electrophysiology and Contractile MediatorsDocument9 pagesControl of Airway Smooth Muscle Tone. I - Electrophysiology and Contractile MediatorsSergio Del AngelNo ratings yet
- Preclinical StudiesDocument16 pagesPreclinical StudiesFadhil Muhammad A.No ratings yet
- Hajos 2000Document11 pagesHajos 2000Breda MarcoNo ratings yet
- Physical Training Reverts Hippocampal Electrophysiological Changes in Rats Submitted To The Pilocarpine Model of EpilepsyDocument7 pagesPhysical Training Reverts Hippocampal Electrophysiological Changes in Rats Submitted To The Pilocarpine Model of EpilepsyJenivia LulileloNo ratings yet
- J Neurophysiol 2000 Xiong 3519 24Document7 pagesJ Neurophysiol 2000 Xiong 3519 24Renato MoraesNo ratings yet
- The Journal of Physiology - 2001 - Naves - Repetitive Nerve Stimulation Decreases The Acetylcholine Content of Quanta atDocument11 pagesThe Journal of Physiology - 2001 - Naves - Repetitive Nerve Stimulation Decreases The Acetylcholine Content of Quanta atAngelita SitumeangNo ratings yet
- Temporal Changes of Post Synaptic Signaling Molecules, Post Synaptic Density-95 and Neuronal Nitric Oxide Synthase, in The Inner Molecular Layer of The Mouse Dentate Gyrus During Voluntary RunningDocument8 pagesTemporal Changes of Post Synaptic Signaling Molecules, Post Synaptic Density-95 and Neuronal Nitric Oxide Synthase, in The Inner Molecular Layer of The Mouse Dentate Gyrus During Voluntary Runningsonjeonggyu87No ratings yet
- Cocaine and HipocampoDocument7 pagesCocaine and HipocampoNico TánatosNo ratings yet
- Dancing nano cell technical data science abh3602_smDocument65 pagesDancing nano cell technical data science abh3602_smbeetlebeeltehocoupleoftimesahNo ratings yet
- Distribution and Ultrastructure of Tyrosine Hydroxylasepositive Neurons in Cns of Bivalve Mollusc Under Action of Elevated Temperature and HypoxiaDocument2 pagesDistribution and Ultrastructure of Tyrosine Hydroxylasepositive Neurons in Cns of Bivalve Mollusc Under Action of Elevated Temperature and HypoxiaBea PippinNo ratings yet
- Gilden Et Al - 1995 - 1-f Noise in Human CognitionDocument4 pagesGilden Et Al - 1995 - 1-f Noise in Human CognitionAli Mert InalNo ratings yet
- Newly Formed Excitatory Pathways Provide A Substrate For Hyperexcitability in Experimental Temporal Lobe EpilepsyDocument12 pagesNewly Formed Excitatory Pathways Provide A Substrate For Hyperexcitability in Experimental Temporal Lobe EpilepsyHsieh Yun JuNo ratings yet
- Cunha-Reis BrJPharm143 (06) 733 (2004)Document12 pagesCunha-Reis BrJPharm143 (06) 733 (2004)dcrrufaNo ratings yet
- European Journal of PharmacologyDocument9 pagesEuropean Journal of PharmacologyMarcelo Pires de OliveiraNo ratings yet
- J. Biol. Chem.-2012-Zhang-33268-81Document15 pagesJ. Biol. Chem.-2012-Zhang-33268-81Soji AdimulaNo ratings yet
- Evaluation of Antiepileptic DrugsDocument32 pagesEvaluation of Antiepileptic DrugsRajpal SinghNo ratings yet
- Diets Prescribed in The Indian SystemDocument20 pagesDiets Prescribed in The Indian SystemBogdan BNo ratings yet
- Alejandra B Cardillo Et Al - Expression of Brugmansia Candida Hyoscyamine 6beta-Hydroxylase Gene in Saccharomyces Cerevisiae and Its Potential Use As BiocatalystDocument7 pagesAlejandra B Cardillo Et Al - Expression of Brugmansia Candida Hyoscyamine 6beta-Hydroxylase Gene in Saccharomyces Cerevisiae and Its Potential Use As BiocatalystFedrmNo ratings yet
- Y. V. Sheludko - Recent Advances in Plant Biotechnology and Genetic Engineering For Production of Secondary MetabolitesDocument9 pagesY. V. Sheludko - Recent Advances in Plant Biotechnology and Genetic Engineering For Production of Secondary MetabolitesFedrmNo ratings yet
- Zhenghao Qian Et Al - A Simple and Efficient Procedure To Enhance Artemisinin Content in Artemisia Annua L. by Seeding To Salinity StressDocument4 pagesZhenghao Qian Et Al - A Simple and Efficient Procedure To Enhance Artemisinin Content in Artemisia Annua L. by Seeding To Salinity StressFedrmNo ratings yet
- Nicholas P. Poolos - The H-Channel: A Potential Channelopathy in Epilepsy?Document6 pagesNicholas P. Poolos - The H-Channel: A Potential Channelopathy in Epilepsy?FedrmNo ratings yet
- Raoufa Abd El-Rahman Et Al - Production of Scopolamine and Hyoscyamine in Calli and Regenerate Cultures of Datura Metel (L.)Document9 pagesRaoufa Abd El-Rahman Et Al - Production of Scopolamine and Hyoscyamine in Calli and Regenerate Cultures of Datura Metel (L.)FedrmNo ratings yet
- Qingming Hou Et Al - Homeostatic Regulation of AMPA Receptor Expression at Single Hippocampal SynapsesDocument6 pagesQingming Hou Et Al - Homeostatic Regulation of AMPA Receptor Expression at Single Hippocampal SynapsesFedrmNo ratings yet
- Javier Palazón Et Al - Application of Metabolic Engineering To The Production of ScopolamineDocument21 pagesJavier Palazón Et Al - Application of Metabolic Engineering To The Production of ScopolamineFedrmNo ratings yet
- Nurussaba Khanam Et Al - Organogenesis, Differentiation and Histolocalization of Alkaloids in Cultured Tissues and Organs of Duboisia Myoporoides R. Br.Document8 pagesNurussaba Khanam Et Al - Organogenesis, Differentiation and Histolocalization of Alkaloids in Cultured Tissues and Organs of Duboisia Myoporoides R. Br.FedrmNo ratings yet
- Minyoung Shin and Dane M. Chetkovich - Activity-Dependent Regulation of H Channel Distribution in Hippocampal CA1 Pyramidal NeuronsDocument13 pagesMinyoung Shin and Dane M. Chetkovich - Activity-Dependent Regulation of H Channel Distribution in Hippocampal CA1 Pyramidal NeuronsFedrmNo ratings yet
- Suvi T. Hakkinen Et Al - Enhanced Secretion of Tropane Alkaloids in Nicotiana Tabacum Hairy Roots Expressing Heterologous Hyoscyamine-6beta-HydroxylaseDocument8 pagesSuvi T. Hakkinen Et Al - Enhanced Secretion of Tropane Alkaloids in Nicotiana Tabacum Hairy Roots Expressing Heterologous Hyoscyamine-6beta-HydroxylaseFedrmNo ratings yet
- Daniel Johnston and Rishikesh Narayanan - Active Dendrites: Colorful Wings of The Mysterious ButterfliesDocument9 pagesDaniel Johnston and Rishikesh Narayanan - Active Dendrites: Colorful Wings of The Mysterious ButterfliesFedrmNo ratings yet
- Rishikesh Narayanan and Daniel Johnston - The H Channel Mediates Location Dependence and Plasticity of Intrinsic Phase Response in Rat Hippocampal NeuronsDocument15 pagesRishikesh Narayanan and Daniel Johnston - The H Channel Mediates Location Dependence and Plasticity of Intrinsic Phase Response in Rat Hippocampal NeuronsFedrmNo ratings yet
- Mariela F. Perez, Francis J. White, and Xiu-Ti Hu - Dopamine D2 Receptor Modulation of K + Channel Activity Regulates Excitability of Nucleus Accumbens Neurons at Different Membrane PotentialsDocument13 pagesMariela F. Perez, Francis J. White, and Xiu-Ti Hu - Dopamine D2 Receptor Modulation of K + Channel Activity Regulates Excitability of Nucleus Accumbens Neurons at Different Membrane PotentialsFedrmNo ratings yet
- Lorenzo A. Cingolani et al- Activity-Dependent Regulation of Synaptic AMPA Receptor Composition and Abundance by β3 IntegrinsDocument30 pagesLorenzo A. Cingolani et al- Activity-Dependent Regulation of Synaptic AMPA Receptor Composition and Abundance by β3 IntegrinsFedrmNo ratings yet
- Martin Biel Et Al - Hyperpolarization-Activated Cation Channels: From Genes To FunctionDocument40 pagesMartin Biel Et Al - Hyperpolarization-Activated Cation Channels: From Genes To FunctionFedrmNo ratings yet
- Neil R. Hardingham Et Al - Presynaptic Efficacy Directs Normalization of Synaptic Strength in Layer 2/3 Rat Neocortex After Paired ActivityDocument12 pagesNeil R. Hardingham Et Al - Presynaptic Efficacy Directs Normalization of Synaptic Strength in Layer 2/3 Rat Neocortex After Paired ActivityFedrmNo ratings yet
- Severine Mahon Et Al - Role of A Striatal Slowly Inactivating Potassium Current in Short-Term Facilitation of Corticostriatal Inputs: A Computer Simulation StudyDocument7 pagesSeverine Mahon Et Al - Role of A Striatal Slowly Inactivating Potassium Current in Short-Term Facilitation of Corticostriatal Inputs: A Computer Simulation StudyFedrmNo ratings yet
- M. Diana Neely Et Al - Cortical Regulation of Dopamine Depletion-Induced Dendritic Spine Loss in Striatal Medium Spiny NeuronsDocument16 pagesM. Diana Neely Et Al - Cortical Regulation of Dopamine Depletion-Induced Dendritic Spine Loss in Striatal Medium Spiny NeuronsFedrmNo ratings yet
- Jason Aoto Et Al - Synaptic Signaling by All-Trans Retinoic Acid in Homeostatic Synaptic PlasticityDocument28 pagesJason Aoto Et Al - Synaptic Signaling by All-Trans Retinoic Acid in Homeostatic Synaptic PlasticityFedrmNo ratings yet
- Jay R. Gibson Et Al - Role For The Subthreshold Currents ILeak and IH in The Homeostatic Control of Excitability in Neocortical Somatostatin-Positive Inhibitory NeuronsDocument15 pagesJay R. Gibson Et Al - Role For The Subthreshold Currents ILeak and IH in The Homeostatic Control of Excitability in Neocortical Somatostatin-Positive Inhibitory NeuronsFedrmNo ratings yet
- Emilie Campanac Et Al - Downregulation of Dendritic Ih in CA1 Pyramidal Neurons After LTPDocument9 pagesEmilie Campanac Et Al - Downregulation of Dendritic Ih in CA1 Pyramidal Neurons After LTPFedrmNo ratings yet
- Claudia Clopath Et Al - Connectivity Reflects Coding: A Model of Voltage-Based Spike-Timing-Dependent-Plasticity With HomeostasisDocument27 pagesClaudia Clopath Et Al - Connectivity Reflects Coding: A Model of Voltage-Based Spike-Timing-Dependent-Plasticity With HomeostasisFedrmNo ratings yet
- Tiago Branco Et Al - Local Dendritic Activity Sets Release Probability at Hippocampal SynapsesDocument11 pagesTiago Branco Et Al - Local Dendritic Activity Sets Release Probability at Hippocampal SynapsesFedrmNo ratings yet
- Clifton C. Rumsey and L. F. Abbott - Synaptic Democracy in Active DendritesDocument13 pagesClifton C. Rumsey and L. F. Abbott - Synaptic Democracy in Active DendritesFedrmNo ratings yet
- Kathleen K. A. Cho Et Al - The Ratio of NR2A/B NMDA Receptor Subunits Determines The Qualities of Ocular Dominance Plasticity in Visual CortexDocument6 pagesKathleen K. A. Cho Et Al - The Ratio of NR2A/B NMDA Receptor Subunits Determines The Qualities of Ocular Dominance Plasticity in Visual CortexFedrmNo ratings yet
- Ithai Rabinowitch and Idan Segev - Two Opposing Plasticity Mechanisms Pulling A Single SynapseDocument7 pagesIthai Rabinowitch and Idan Segev - Two Opposing Plasticity Mechanisms Pulling A Single SynapseFedrmNo ratings yet
- Ingrid Van Welie Et Al - Different Levels of Ih Determine Distinct Temporal Integration in Bursting and Regular-Spiking Neurons in Rat SubiculumDocument12 pagesIngrid Van Welie Et Al - Different Levels of Ih Determine Distinct Temporal Integration in Bursting and Regular-Spiking Neurons in Rat SubiculumFedrmNo ratings yet
- Peter A. Appleby and Terry Elliott - Synaptic and Temporal Ensemble Interpretation of Spike-Timing-Dependent PlasticityDocument21 pagesPeter A. Appleby and Terry Elliott - Synaptic and Temporal Ensemble Interpretation of Spike-Timing-Dependent PlasticityFedrmNo ratings yet
- Jonathan E Rubin - Steady States in An Iterative Model For Multiplicative Spike-Timing-Dependent PlasticityDocument10 pagesJonathan E Rubin - Steady States in An Iterative Model For Multiplicative Spike-Timing-Dependent PlasticityFedrmNo ratings yet
- Ecg Stemi: Sequence of Changes in Evolving STEMIDocument3 pagesEcg Stemi: Sequence of Changes in Evolving STEMIFajar AgNo ratings yet
- C3 Carbon Fixation: Key Photosynthesis ProcessDocument8 pagesC3 Carbon Fixation: Key Photosynthesis ProcessBashiir NuurNo ratings yet
- Biology Assignment 2 Atefah Razack 1Document15 pagesBiology Assignment 2 Atefah Razack 1api-491258706No ratings yet
- What Is The Rate For Performing Chest Compressions For A Victim of Any Age A-30 Compressions Per Minute B - 50 Compressions Per Minute C - 80 Compressions Per MinuteDocument7 pagesWhat Is The Rate For Performing Chest Compressions For A Victim of Any Age A-30 Compressions Per Minute B - 50 Compressions Per Minute C - 80 Compressions Per MinuteHassan Shehri100% (2)
- Junctional: TachycardiaDocument11 pagesJunctional: TachycardiachaiNo ratings yet
- Respiratory Distress Syndrome (Hyaline Membrane Disease)Document98 pagesRespiratory Distress Syndrome (Hyaline Membrane Disease)Miraf MesfinNo ratings yet
- Calculation of q10 Value-1Document2 pagesCalculation of q10 Value-1Leighton ThompsonNo ratings yet
- Effects of A Balloon-Blowing Exercise On Lung Function of SmokersDocument4 pagesEffects of A Balloon-Blowing Exercise On Lung Function of SmokersMónica ReisNo ratings yet
- Lesson 1 - Introduction To AnemiaDocument10 pagesLesson 1 - Introduction To AnemiaThobby GamosaNo ratings yet
- Biology worksheet answersDocument2 pagesBiology worksheet answersAziyaNo ratings yet
- Human Body WebQuestDocument6 pagesHuman Body WebQuestjennyferchoNo ratings yet
- Q2 STEM General Biology 1 Week 2Document4 pagesQ2 STEM General Biology 1 Week 2MarkElijah ObelNo ratings yet
- PROTOCOL of Management of Critical CasesDocument131 pagesPROTOCOL of Management of Critical Casesaziz.shokry.mosaNo ratings yet
- Pulm 2005 Exam QuestionsDocument32 pagesPulm 2005 Exam QuestionsItharshan IndreswaranNo ratings yet
- Pediatric Critical Care Nutrition PDFDocument324 pagesPediatric Critical Care Nutrition PDFNia DefinisiNo ratings yet
- FINAL Lab Report #3Document10 pagesFINAL Lab Report #3Chippy RabeNo ratings yet
- TUGAS BAHASA INGGRIS 2 (Devi Yuliyanti)Document4 pagesTUGAS BAHASA INGGRIS 2 (Devi Yuliyanti)DeviNo ratings yet
- Blood ComponentsDocument2 pagesBlood ComponentsParul ShahNo ratings yet
- Lecture - 5 PDFDocument5 pagesLecture - 5 PDFDavid JokerNo ratings yet
- Anesthetic Considerations For Geriatric DogsDocument3 pagesAnesthetic Considerations For Geriatric DogsFernanda PérezNo ratings yet
- ABG Analysis: 6-Step ApproachDocument22 pagesABG Analysis: 6-Step Approachrajan40dmcNo ratings yet
- (Pharm) 1s-2 Ans DrugsDocument16 pages(Pharm) 1s-2 Ans DrugsKim Ramos67% (3)
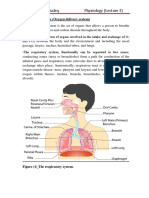
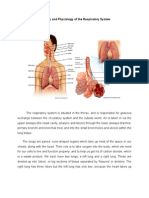
- Anatomy and Physiology of The Respiratory SystemDocument4 pagesAnatomy and Physiology of The Respiratory SystemchacameroNo ratings yet
- Pulmonary Function TestingDocument3 pagesPulmonary Function TestingAditya KumarNo ratings yet
- Pediatric Hematology, 3rd Ed. 2006Document2 pagesPediatric Hematology, 3rd Ed. 2006Henny HansengNo ratings yet
- The Utility of Non-Invasive Brain Stimulation in RDocument10 pagesThe Utility of Non-Invasive Brain Stimulation in RsanagaumiNo ratings yet
- Worksheet - Respiratory System PDFDocument3 pagesWorksheet - Respiratory System PDFDezta Fandity' VantycaNo ratings yet
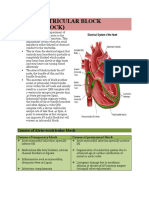
- Atrioventricular Block (Heart Block) : Causes of Atrio-Ventricular BlockDocument10 pagesAtrioventricular Block (Heart Block) : Causes of Atrio-Ventricular BlockLiza M. PurocNo ratings yet
- Laboratory Manual For Human Anatomy and Physiology Main Version 4th Edition Martin Solutions ManualDocument11 pagesLaboratory Manual For Human Anatomy and Physiology Main Version 4th Edition Martin Solutions ManualBradSalazarogzn100% (16)
- Drug Study: Medication Indication Contraindicati0N Side Effects Use Caution inDocument17 pagesDrug Study: Medication Indication Contraindicati0N Side Effects Use Caution inAngely Dianne Santiago II100% (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet