You might also like
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Clase005 Manchas Bioquímica ForenseDocument64 pagesClase005 Manchas Bioquímica Forenseapi-371018683% (6)
- Guía003 - Organización Subcelular - Biología Celular Lab OratorioDocument11 pagesGuía003 - Organización Subcelular - Biología Celular Lab Oratorioapi-3710186100% (7)
- Trabajo Grupal - Segunda Unidad - Resistencia de MaterialesDocument12 pagesTrabajo Grupal - Segunda Unidad - Resistencia de MaterialesChristian Jair Sandoval Damián50% (2)
- Clase Región Pelvis AnatomiaDocument29 pagesClase Región Pelvis Anatomiaapi-3707147100% (2)
- Exposición - Síndrome Vangierke - Introducción A Bioquímica IIDocument17 pagesExposición - Síndrome Vangierke - Introducción A Bioquímica IIapi-3710186100% (2)
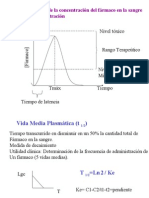
- Clase002 Vida Media FarmacodinámiaDocument47 pagesClase002 Vida Media Farmacodinámiaapi-3710186100% (1)
- Clase001 Introducción FarmacodinámiaDocument66 pagesClase001 Introducción Farmacodinámiaapi-3710186No ratings yet
- Exposición - VIH - Introducción A Bioquímica IIDocument11 pagesExposición - VIH - Introducción A Bioquímica IIapi-3710186100% (1)
- Exposición - Pelagra - Introducción A Bioquímica IIDocument21 pagesExposición - Pelagra - Introducción A Bioquímica IIapi-3710186No ratings yet
- Exposición - Porfíria - Introducción A Bioquímica IIDocument12 pagesExposición - Porfíria - Introducción A Bioquímica IIapi-3710186No ratings yet
- Exposición - Priones - Introducción A Bioquímica IIDocument14 pagesExposición - Priones - Introducción A Bioquímica IIapi-3710186100% (1)
- Exposición - Intolerancia A La Lactosa - Introducción A Bioquímica IIDocument11 pagesExposición - Intolerancia A La Lactosa - Introducción A Bioquímica IIapi-3710186No ratings yet
- Exposición - Colágeno - Introducción A Bioquímica IIDocument12 pagesExposición - Colágeno - Introducción A Bioquímica IIapi-3710186No ratings yet
- Exposición - Catarátas - Introducción A Bioquímica IIDocument17 pagesExposición - Catarátas - Introducción A Bioquímica IIapi-3710186No ratings yet
- c9f6745ebf680e99fc97e8d06b70bb40 (2).docxDocument7 pagesc9f6745ebf680e99fc97e8d06b70bb40 (2).docxJohn Jairo Vides IribeNo ratings yet
- PVPP Concepcion PDFDocument292 pagesPVPP Concepcion PDFJuan Carlos Sedano LunaNo ratings yet
- Combinatoria UltimoDocument23 pagesCombinatoria UltimoAlex Jhonny Crispin FernandezNo ratings yet
- Abscesos ApicalesDocument4 pagesAbscesos Apicalesjoancys guevaraNo ratings yet
- Congestion de RedDocument26 pagesCongestion de RedLeonardo Serna GuarínNo ratings yet
- 1ro Experiencia de Aprendizaje #4.Document21 pages1ro Experiencia de Aprendizaje #4.Dannis Nayeli Manrique ChecaNo ratings yet
- Revista Tecnologia Minera 41Document271 pagesRevista Tecnologia Minera 41Manuel AlonsoNo ratings yet
- Aguila Sa de CVDocument3 pagesAguila Sa de CVPablo Hernandez TequianesNo ratings yet
- Sergio MendezDocument4 pagesSergio MendezFredi CastilloNo ratings yet
- Copia de CASO PRÁCTICO DESARROLLO EMPRENDEDORDocument7 pagesCopia de CASO PRÁCTICO DESARROLLO EMPRENDEDORCadena Sánchez María FernandaNo ratings yet
- GAGM - Edison David Arias y Victor Manuel RodríguezDocument78 pagesGAGM - Edison David Arias y Victor Manuel Rodríguezvictor_147No ratings yet
- Metodos IIDocument4 pagesMetodos IINehemias Lopez100% (1)
- Actividad de Aprendizaje #01: Reflexionando Sobre El Medio Ambiente A Través Del ArteDocument3 pagesActividad de Aprendizaje #01: Reflexionando Sobre El Medio Ambiente A Través Del ArteJOSEPH VALENTINO GOMEZ BENITESNo ratings yet
- Taller 2 ContabilidadDocument4 pagesTaller 2 ContabilidadenchozxNo ratings yet
- Manual de Fundamento de La MiliciaDocument68 pagesManual de Fundamento de La MiliciaCinunefaupata Hipódromo72% (32)
- Patología VeterinariaDocument14 pagesPatología VeterinariaCatherin EstupiñanNo ratings yet
- Cadena Productiva Del MangoDocument27 pagesCadena Productiva Del MangoYirley MarínNo ratings yet
- MAZAMORRADocument7 pagesMAZAMORRAVP FiorelaNo ratings yet
- Informe Carolina 1 (2) INFORME EVALUA - 10Document10 pagesInforme Carolina 1 (2) INFORME EVALUA - 10Yarii SandovalNo ratings yet
- Condensador en Serie y Paralelo en CCDocument6 pagesCondensador en Serie y Paralelo en CCMajo SalgadoNo ratings yet
- PasteleriaDocument12 pagesPasteleriaNicolas SuhNo ratings yet
- MANUAL FUSION 3 EspañolDocument32 pagesMANUAL FUSION 3 EspañolVicente Luna GarcíaNo ratings yet
- Actividad 2 - Tema 1 AngelDocument4 pagesActividad 2 - Tema 1 AngelAngel LopezNo ratings yet
- Preguntas Teóricas - Licencia en Pmá.Document5 pagesPreguntas Teóricas - Licencia en Pmá.bigghost030% (1)
- Tema 11 La Administracion de La EmpresaDocument3 pagesTema 11 La Administracion de La EmpresaSusanaNo ratings yet
- Anexo 1 Rubrica Examen de Grado 2021-2022Document5 pagesAnexo 1 Rubrica Examen de Grado 2021-2022Kerly Yaselga100% (1)
- Proyecto La Floresta 2 - Grupo 1Document32 pagesProyecto La Floresta 2 - Grupo 1Alejandro CanchalaNo ratings yet
- Proceso de Proyeccion SocialDocument4 pagesProceso de Proyeccion Socialhernando luis sanchez aguirreNo ratings yet
- Ensayo CriticoDocument3 pagesEnsayo Criticojoel jesus avalos zamudioNo ratings yet