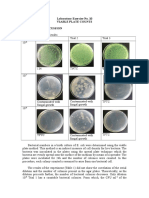
You might also like
- CH 12 Instructor GuideDocument11 pagesCH 12 Instructor GuideVee Nara0% (1)
- Sargassum MuticumDocument10 pagesSargassum MuticumtzintzuntzinNo ratings yet
- Industrial Production of Metabolites - Organic Acids, Alcohols, AntibioticsDocument17 pagesIndustrial Production of Metabolites - Organic Acids, Alcohols, Antibioticsvaibhav_ta0% (1)
- Heterotrophic Planktonic Microbes: Virus, Bacteria, Archaea, and ProtozoaDocument34 pagesHeterotrophic Planktonic Microbes: Virus, Bacteria, Archaea, and ProtozoaCharity MbiziNo ratings yet
- Microbial EcologyDocument30 pagesMicrobial EcologyRavi Sharma100% (1)
- Microbiology and MycologyDocument76 pagesMicrobiology and MycologyAhmad Said AliNo ratings yet
- TCBS Agar PDFDocument2 pagesTCBS Agar PDFJhudit Camacho100% (1)
- 6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthDocument6 pages6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthHanief MitsukhaNo ratings yet
- IntroductionDocument91 pagesIntroductionmulatumelese100% (1)
- FungusDocument31 pagesFungushadiya rehmanNo ratings yet
- New Developments in Chromogenic and Fluorogenic Culture MediaDocument14 pagesNew Developments in Chromogenic and Fluorogenic Culture MediaPaulina LizzethNo ratings yet
- University Malaysia Sabah - Lab 2Document10 pagesUniversity Malaysia Sabah - Lab 2Nur Nadiah Kamaruzaman50% (2)
- Strategies For Culture of Unculturable BacteriaDocument7 pagesStrategies For Culture of Unculturable BacteriaEd M. K-zhan NözNo ratings yet
- Module 1 - Introduction To MicrobiologyDocument14 pagesModule 1 - Introduction To MicrobiologyRealyn FababaerNo ratings yet
- FST5206 - 2018/2019-1 (Microbiological Food Safety) Lecture 1. Microbial Ecology 1Document97 pagesFST5206 - 2018/2019-1 (Microbiological Food Safety) Lecture 1. Microbial Ecology 1Nurul NaqibahNo ratings yet
- Secondary MetabolliteDocument20 pagesSecondary MetabolliteChindieyciiEy Lebaiey CwekZrdoghNo ratings yet
- Module 4 Annex Part 1 MKDocument95 pagesModule 4 Annex Part 1 MKtata_77No ratings yet
- Salmonella - Methods and Protocols.-springer-Verlag New York (2020)Document215 pagesSalmonella - Methods and Protocols.-springer-Verlag New York (2020)Yudith ParamitadeviNo ratings yet
- Guide To The Classification and Identification of The Actinomycetes and Their Antibiotics by Waksman, Selman A. (Selman Abraham), 1888-1973 Lechevalier, Hubert ADocument264 pagesGuide To The Classification and Identification of The Actinomycetes and Their Antibiotics by Waksman, Selman A. (Selman Abraham), 1888-1973 Lechevalier, Hubert AVictor R SatrianiNo ratings yet
- Fungi PDFDocument64 pagesFungi PDFSilvia Tri ayuNo ratings yet
- Isolation, Identification, & Preservation of IndusrtialDocument16 pagesIsolation, Identification, & Preservation of IndusrtialPranav Nakhate100% (1)
- Use of Micropippettor and SpectrophotometerDocument6 pagesUse of Micropippettor and SpectrophotometerMichelleNo ratings yet
- Dyes and StainsDocument49 pagesDyes and StainsBilal Mumtaz AwanNo ratings yet
- Phylogenetic TreesDocument20 pagesPhylogenetic TreesRamesh Kandagatla100% (1)
- PN ChromogenicMediaDocument12 pagesPN ChromogenicMediaNguyen Ngoc ThachNo ratings yet
- Branches of MicrobiologyDocument2 pagesBranches of Microbiologyhaseeb_khan321294067% (3)
- Bio Sketch SampleDocument4 pagesBio Sketch SampleLuis Emilio RosarioNo ratings yet
- (Advances in Parasitology 76) Louis M. Weiss and Herbert B. Tanowitz (Eds.) - Chagas Disease, Part B-Academic Press, Elsevier (2011)Document283 pages(Advances in Parasitology 76) Louis M. Weiss and Herbert B. Tanowitz (Eds.) - Chagas Disease, Part B-Academic Press, Elsevier (2011)Stoian GoranovNo ratings yet
- DocumentDocument15 pagesDocumentJennifer BanteNo ratings yet
- Scope of MicrobiologyDocument9 pagesScope of MicrobiologyrachnnaaNo ratings yet
- HookwormDocument9 pagesHookwormfadilah mutiaNo ratings yet
- Miller/Tupper: Zoology 11e Instructor's Manual: Chapter SummaryDocument3 pagesMiller/Tupper: Zoology 11e Instructor's Manual: Chapter SummaryCaesar Franz RuizNo ratings yet
- Classification and Indentification of BacteriaDocument77 pagesClassification and Indentification of BacteriaAnand MadhavanNo ratings yet
- Escherichia Coli and Total Coliform Using The IDEXX Quanti-Tray 2000 System With Colilert ReagentDocument17 pagesEscherichia Coli and Total Coliform Using The IDEXX Quanti-Tray 2000 System With Colilert ReagentchristianNo ratings yet
- Laboratory ManualDocument32 pagesLaboratory ManuallkjwrotuNo ratings yet
- 01 Introduction To MicrobiologyDocument46 pages01 Introduction To MicrobiologyAnabel ManggagaNo ratings yet
- Tested Studies For Laboratory Teaching. Peifer. 1989Document174 pagesTested Studies For Laboratory Teaching. Peifer. 1989NidzaNo ratings yet
- Fungi: Morphology and ClassificationDocument37 pagesFungi: Morphology and Classificationjihan faradhilaNo ratings yet
- Methods To Study Soil Microbial DiversityDocument25 pagesMethods To Study Soil Microbial Diversityrd2165scribdNo ratings yet
- Zoology ManualDocument85 pagesZoology ManualShareena M R. EpresNo ratings yet
- Alternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesDocument4 pagesAlternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesMasitha Dwi AmiraNo ratings yet
- In Vitro Methods For Detecting Cytotoxicity: UNIT 2.6Document27 pagesIn Vitro Methods For Detecting Cytotoxicity: UNIT 2.6hiền nguyễnNo ratings yet
- Bacterial ClassificationDocument62 pagesBacterial ClassificationQuan ThieuNo ratings yet
- Laboratory Exercise No. 10 Viable Plate Counts Results and DiscussionDocument3 pagesLaboratory Exercise No. 10 Viable Plate Counts Results and Discussionvanessa olga100% (1)
- FTM 299417Document7 pagesFTM 299417Chia-Yi HsuNo ratings yet
- Biodiversity of Cyanobacteria and Green Algae On MonumentsDocument15 pagesBiodiversity of Cyanobacteria and Green Algae On MonumentsWendy PhNo ratings yet
- Bacterial GrowthDocument5 pagesBacterial GrowthVytheeshwaran Vedagiri100% (20)
- General Microbiology Laboratory Manual: Biology 490Document83 pagesGeneral Microbiology Laboratory Manual: Biology 490Ibnul MubarokNo ratings yet
- General Microbiology (Chapter 3)Document18 pagesGeneral Microbiology (Chapter 3)Ashraf OsmanNo ratings yet
- 3.1 Introduction To Fungi and MycosesDocument26 pages3.1 Introduction To Fungi and MycosesJoshua KalundaNo ratings yet
- Comparison of Methods Used For Assessing The Viability and Vitality of Yeast CellsDocument12 pagesComparison of Methods Used For Assessing The Viability and Vitality of Yeast CellsKothari VishalNo ratings yet
- Moving Environmental DNA Methods From Concept To Practice For Monitoring Aquatic MacroorganismsDocument3 pagesMoving Environmental DNA Methods From Concept To Practice For Monitoring Aquatic MacroorganismsFrancisco PrietoNo ratings yet
- Principles and Types of StainingDocument42 pagesPrinciples and Types of StainingMthandeni KhumaloNo ratings yet
- Prokaryote NomenclatureDocument6 pagesProkaryote NomenclatureEnrique E. ZuñigaNo ratings yet
- HSC Biotech OptionDocument3 pagesHSC Biotech OptionthalwiNo ratings yet
- Rapid Methods For The Detection of Foodborne Bacterial PathogensDocument19 pagesRapid Methods For The Detection of Foodborne Bacterial PathogensIoana MoldovanNo ratings yet
- 5 Classification of MicroorganismsDocument4 pages5 Classification of MicroorganismsMaeve Ylain SeanNo ratings yet
- Biochemical Tests For BacteriaDocument16 pagesBiochemical Tests For BacteriaNichols Amy Tarun100% (1)
- BT1008 Microbiology LessonplanDocument2 pagesBT1008 Microbiology Lessonplansangeeta palNo ratings yet
- Rapid Detection and Identification of Infectious AgentsFrom EverandRapid Detection and Identification of Infectious AgentsDavid KingsburyNo ratings yet
- Steps To Be Followed To Rename or Move Control Files LocationDocument8 pagesSteps To Be Followed To Rename or Move Control Files Locationvaibhav_taNo ratings yet
- SAP Line OpenerDocument10 pagesSAP Line Openervaibhav_taNo ratings yet
- Step by Step Procedure For SSO Between EP PDFDocument10 pagesStep by Step Procedure For SSO Between EP PDFvaibhav_taNo ratings yet
- Bureau of Indian Standards PDFDocument24 pagesBureau of Indian Standards PDFvaibhav_taNo ratings yet
- Step by Step Guide On Creating Physical Standby Using RMAN (ASM) - Oracle Database Administration PDFDocument13 pagesStep by Step Guide On Creating Physical Standby Using RMAN (ASM) - Oracle Database Administration PDFvaibhav_taNo ratings yet
- BRTools To Backup SAP On ORACLEDocument12 pagesBRTools To Backup SAP On ORACLEOthman Hendy Suseno100% (1)
- Biofertilizer ManualDocument138 pagesBiofertilizer Manualvaibhav_taNo ratings yet
- Biological Forum-An International Journal, Spl. Iss. 4 (1) (2012)Document237 pagesBiological Forum-An International Journal, Spl. Iss. 4 (1) (2012)vaibhav_ta67% (3)
- Environmental Science Literature Review ExampleDocument5 pagesEnvironmental Science Literature Review Exampledajemevefaz2No ratings yet
- SCI403 - GRP 1 - 3MBIO6 - Change of Thesis FormDocument5 pagesSCI403 - GRP 1 - 3MBIO6 - Change of Thesis FormFert FreNo ratings yet
- ADM - Grade 11 - Quarter1 (S1112ES-Ia-e-6)Document24 pagesADM - Grade 11 - Quarter1 (S1112ES-Ia-e-6)josephine alcantaraNo ratings yet
- Tizon Let's Check#3Document2 pagesTizon Let's Check#3Dale TizonNo ratings yet
- Sample Data and Problems Project For HydrologyDocument42 pagesSample Data and Problems Project For HydrologyEugene CardamaNo ratings yet
- How Does The 4Rs Framework Guide Deeper Reflection in STAR-L/P?Document1 pageHow Does The 4Rs Framework Guide Deeper Reflection in STAR-L/P?shivani gargNo ratings yet
- Modelling The Structure of The Earth - Student Activity-1Document5 pagesModelling The Structure of The Earth - Student Activity-1Macy RiegoNo ratings yet
- Phenomenon of Experiential Space: by Shruti GuptaDocument4 pagesPhenomenon of Experiential Space: by Shruti GuptaPrerana CholakheNo ratings yet
- Geology Assignment by MmsDocument8 pagesGeology Assignment by MmsMaddyNo ratings yet
- EST MicroprojectDocument12 pagesEST MicroprojectRohan ChavanNo ratings yet
- E.P.C 321 2014Document1,003 pagesE.P.C 321 2014Mich KidNo ratings yet
- Bioreceptivity - WikipediaDocument3 pagesBioreceptivity - Wikipediaapi-690823987No ratings yet
- ColiDocument6 pagesColimaryanaNo ratings yet
- Science Module Quarter 4 L1Document4 pagesScience Module Quarter 4 L1Alex Tutor100% (1)
- AV3Document5 pagesAV3nguyenkimngan0806No ratings yet
- Physics LadakhDocument10 pagesPhysics LadakhAdarsha NNo ratings yet
- EcotoneDocument10 pagesEcotonePrashant KumarNo ratings yet
- Chapter 1Document4 pagesChapter 1Xylafe MonetteNo ratings yet
- (English (Auto-Generated) ) I Thought Plastic Pollution Wasn't My Problem ... - Archie Wallyn - TEDxYouth@DPL (DownSub - Com)Document3 pages(English (Auto-Generated) ) I Thought Plastic Pollution Wasn't My Problem ... - Archie Wallyn - TEDxYouth@DPL (DownSub - Com)Via ResvianaNo ratings yet
- Environmental Impact Assessment and Management PlanDocument6 pagesEnvironmental Impact Assessment and Management PlanWael AlmahdiNo ratings yet
- Appendix G Construction Inspection Checklists and Asbuilt CertificationsDocument30 pagesAppendix G Construction Inspection Checklists and Asbuilt CertificationsMehdi RezougNo ratings yet
- College of Agricultural Science: Course Code Hort - 381Document39 pagesCollege of Agricultural Science: Course Code Hort - 381TOLA ABENo ratings yet
- Revenue Sharing Coordination in Reverse LogisticsDocument12 pagesRevenue Sharing Coordination in Reverse LogisticsKatarina da Silva CoutoNo ratings yet
- 7 АНГИDocument10 pages7 АНГИEnglish EduNo ratings yet
- Smile Cesc Q1 LP 3Document10 pagesSmile Cesc Q1 LP 3Jonalyn TandaanNo ratings yet
- Causes and Consequences of Spoliation in The City of Kinshasa, The Democratic Republic of The CongoDocument4 pagesCauses and Consequences of Spoliation in The City of Kinshasa, The Democratic Republic of The CongoInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Bakir 2006 Policy Agenda Setting and Risk CommunicationDocument22 pagesBakir 2006 Policy Agenda Setting and Risk Communicationaryani nabillaNo ratings yet
- REVIEW 251-500 Ans.Document60 pagesREVIEW 251-500 Ans.patelrohil2006No ratings yet
- De Minh HoaDocument6 pagesDe Minh Hoahuycao6dNo ratings yet
- Background of Manjung Municipal CouncilDocument3 pagesBackground of Manjung Municipal CouncilaishahNo ratings yet