You might also like
- ABB 3wdg TRAFO Data SheetDocument2 pagesABB 3wdg TRAFO Data SheetKunik SwaroopNo ratings yet
- Generational CursesDocument5 pagesGenerational Cursesfatima110No ratings yet
- Numerical Reasoning FormulasDocument7 pagesNumerical Reasoning FormulasRachid HC100% (1)
- FOOD AND NUTRITIONAL ANALYSIS Fruits and Fruit Products-334-341Document8 pagesFOOD AND NUTRITIONAL ANALYSIS Fruits and Fruit Products-334-341fernandoferozNo ratings yet
- Changes of Isoflavones During Processing of Soy Protein IsolatesDocument5 pagesChanges of Isoflavones During Processing of Soy Protein IsolatesDinh DungNo ratings yet
- Vasco 2008 Food-ChemistryDocument8 pagesVasco 2008 Food-ChemistryzenufanaNo ratings yet
- Food Research International: Alma Fernanda Sánchez Maldonado, Elizabeth Mudge, Michael G. Gänzle, Andreas SchieberDocument8 pagesFood Research International: Alma Fernanda Sánchez Maldonado, Elizabeth Mudge, Michael G. Gänzle, Andreas SchieberMaria MihaelaNo ratings yet
- Buffalo Cheese Whey Hydrolyzed With Alcalase As An AntibrowningDocument8 pagesBuffalo Cheese Whey Hydrolyzed With Alcalase As An Antibrowningdea wulanNo ratings yet
- Analysis and Identification of Phenolic Compounds in Dioscorea Hispida DennstDocument14 pagesAnalysis and Identification of Phenolic Compounds in Dioscorea Hispida DennstRainolNo ratings yet
- Polar Lipids: Biology, Chemistry, and TechnologyFrom EverandPolar Lipids: Biology, Chemistry, and TechnologyMoghis U. AhmadNo ratings yet
- Effect of Coating and Wrapping Materials On The Shelf Life of Apple Malus Domestica CV BorkhDocument11 pagesEffect of Coating and Wrapping Materials On The Shelf Life of Apple Malus Domestica CV BorkhNadia Aprili PutriNo ratings yet
- J Agric Food Chem 2005,53,1370Document4 pagesJ Agric Food Chem 2005,53,1370Teo SuazoNo ratings yet
- Molecules AlcaparrasDocument16 pagesMolecules AlcaparrasJosé Vasquez SucreNo ratings yet
- Potential of Lemongrass Leaves Extract Cymbopogon Citratus As Prevention For Oil OxidationDocument6 pagesPotential of Lemongrass Leaves Extract Cymbopogon Citratus As Prevention For Oil OxidationAdriana Lucia MerchanNo ratings yet
- Full Text 6 - Flavonoids From Almond Skins Are Bioavailable and Act SynergisticallyDocument8 pagesFull Text 6 - Flavonoids From Almond Skins Are Bioavailable and Act SynergisticallyPencari IlmuNo ratings yet
- Effects of Dietary Supplementation With Sea Buckthorn (Hippophae Atopic DermatitisDocument9 pagesEffects of Dietary Supplementation With Sea Buckthorn (Hippophae Atopic DermatitiswxcvbnnbvcxwNo ratings yet
- 47 Oprescu e 9 19Document4 pages47 Oprescu e 9 19Sandy Wahyu DNo ratings yet
- 9.kanlayavattanakul, M., Lourith, N.Document8 pages9.kanlayavattanakul, M., Lourith, N.27. SYAHRIN AZRIL IZULKHAQUENo ratings yet
- E fficient Separation and Analysis of Triacylglycerols: Quantitation of β‑Palmitate (OPO) in Oils and Infant FormulasDocument8 pagesE fficient Separation and Analysis of Triacylglycerols: Quantitation of β‑Palmitate (OPO) in Oils and Infant FormulasReinaldo Ongky Billy AnandoNo ratings yet
- Antioxidants 03 00038Document17 pagesAntioxidants 03 00038Diệu LinhNo ratings yet
- TFC RPCDocument9 pagesTFC RPCPhtthiphong chansingeonNo ratings yet
- N Fructo OligosaccharidesDocument5 pagesN Fructo Oligosaccharidesphuongdung2012No ratings yet
- Characterization of Bioactive Compounds From Monovarietal Virgin Olive Oils Relationship Between Phenolic Compounds Antioxidant CapacitiesDocument12 pagesCharacterization of Bioactive Compounds From Monovarietal Virgin Olive Oils Relationship Between Phenolic Compounds Antioxidant CapacitiesDina BougandouraNo ratings yet
- Anticancer Article 7Document14 pagesAnticancer Article 720-20311-014No ratings yet
- Amino Acid 3Document6 pagesAmino Acid 3sarita choudharyNo ratings yet
- Foods: Production of Bio-Functional Protein Through Revalorization of Apricot Kernel CakeDocument12 pagesFoods: Production of Bio-Functional Protein Through Revalorization of Apricot Kernel CakePenelope MeloNo ratings yet
- Salfarina RamliDocument9 pagesSalfarina RamliMellisa Laura MintoroNo ratings yet
- Research Project Burch Micheal - 2Document44 pagesResearch Project Burch Micheal - 2Bless UgbongNo ratings yet
- Odbayar 2006Document5 pagesOdbayar 2006Corona FingerNo ratings yet
- Essential Chemicals in Selected Fruit Peels From Manila, PhilippinesDocument5 pagesEssential Chemicals in Selected Fruit Peels From Manila, PhilippinesPeterson RonquilloNo ratings yet
- Dragon FruitDocument8 pagesDragon FruitLive LikeNo ratings yet
- Avocado Seeds: Extraction Optimization and Possible Use As Antioxidant in FoodDocument30 pagesAvocado Seeds: Extraction Optimization and Possible Use As Antioxidant in FoodĐeŋýzař Đ ZòįđýckNo ratings yet
- Functionality and Antioxidant Properties of Tilapia (Oreochromis Niloticus) As Influenced by The Degree of HydrolysisDocument19 pagesFunctionality and Antioxidant Properties of Tilapia (Oreochromis Niloticus) As Influenced by The Degree of HydrolysisasepawaNo ratings yet
- Patulin Analysis With LC-MSDocument5 pagesPatulin Analysis With LC-MSbohemian84No ratings yet
- And Fermented Catfish Oil)Document9 pagesAnd Fermented Catfish Oil)Ikcal SaputraNo ratings yet
- OJC Vol34 No2 P 1136-1140Document5 pagesOJC Vol34 No2 P 1136-1140be21b002No ratings yet
- Exposure To Patulin From Consumption of Apple Based ProductsDocument8 pagesExposure To Patulin From Consumption of Apple Based ProductsSilviaBernalNo ratings yet
- Minitab17 GettingStarted enDocument6 pagesMinitab17 GettingStarted enTan Hau VoNo ratings yet
- 2019-JDS 16729Document14 pages2019-JDS 16729Miguel Fernandez NiñoNo ratings yet
- 09e41508ace8b77161000000 PDFDocument7 pages09e41508ace8b77161000000 PDFNandika Kasaru JackNo ratings yet
- A Lignocellulolytic Enzyme System For FRDocument241 pagesA Lignocellulolytic Enzyme System For FRSachin SharmaNo ratings yet
- Potency of CitrusDocument9 pagesPotency of CitrusYulitha Rempe SampeallaNo ratings yet
- Cellulolytic Fruits Wastes: A Potential Support For Enzyme Assisted Protein ProductionDocument9 pagesCellulolytic Fruits Wastes: A Potential Support For Enzyme Assisted Protein ProductionkesavarajNo ratings yet
- Cymbopogon Citratus Leaves Characterization of Flavonoids by HPLC PDA ESI MS MS and An Approach To Their Potential As A Source of Bioactive PolyphenolDocument11 pagesCymbopogon Citratus Leaves Characterization of Flavonoids by HPLC PDA ESI MS MS and An Approach To Their Potential As A Source of Bioactive PolyphenolGabriel Oliveira de SouzaNo ratings yet
- Cosmo SurfaceDocument24 pagesCosmo SurfaceDhiraviya DeviNo ratings yet
- Characterization and Application of Gelatin FilmsDocument23 pagesCharacterization and Application of Gelatin FilmsBETÜL KILINÇLINo ratings yet
- FNS 2015052610384538Document9 pagesFNS 2015052610384538Sayuri hernandez vargasNo ratings yet
- Artículo 2Document14 pagesArtículo 2Frida LiraNo ratings yet
- Research Article Spirulina: Quantification of Phytochemicals From Commercial Products and Their Antioxidant ActivitiesDocument14 pagesResearch Article Spirulina: Quantification of Phytochemicals From Commercial Products and Their Antioxidant ActivitiesGouriJayanNo ratings yet
- Scientific Paper On Determination of Reducing Sugars in Fruit and VegetablesDocument3 pagesScientific Paper On Determination of Reducing Sugars in Fruit and VegetablesJulius BelarmaNo ratings yet
- Studies On Indian Green Leafy Vegetables For Their Antioxidant ActivityDocument7 pagesStudies On Indian Green Leafy Vegetables For Their Antioxidant ActivityAlcides SitoeNo ratings yet
- Lycopene Concentration and Physico-Chemical Properties of Tropical FruitsDocument5 pagesLycopene Concentration and Physico-Chemical Properties of Tropical FruitsSanTilinNo ratings yet
- Acidos Grasos SIDocument7 pagesAcidos Grasos SIAlyssa Li SalazarNo ratings yet
- Mahayothee 2016Document12 pagesMahayothee 2016Fauzi SatriaNo ratings yet
- J 0814248Document7 pagesJ 0814248Trinidad FigueroaNo ratings yet
- 5 - Structural and Functional Characterization of Polyphenols Isolated From Acerola Malpighia Emarginata DC FruitDocument8 pages5 - Structural and Functional Characterization of Polyphenols Isolated From Acerola Malpighia Emarginata DC FruitTALITA TALITANo ratings yet
- Nutrients: Extracts, Anthocyanins and Procyanidins From Enzyme InhibitorsDocument16 pagesNutrients: Extracts, Anthocyanins and Procyanidins From Enzyme InhibitorsLavanya Priya SathyanNo ratings yet
- PenelitianDocument6 pagesPenelitianAdrin Ma'rufNo ratings yet
- In Vitro Callus and in Vivo Leaf Extract of Gymnema Sylvestre Stimulate - Cells Regeneration and Anti-Diabetic Activity in Wistar RatsDocument7 pagesIn Vitro Callus and in Vivo Leaf Extract of Gymnema Sylvestre Stimulate - Cells Regeneration and Anti-Diabetic Activity in Wistar Ratsapi-198310771No ratings yet
- Effects of Processing On Physicochemical and Antinutritional Properties of Black Turtle Bean (Phaseolus Vulgaris L.) Seeds FlourDocument11 pagesEffects of Processing On Physicochemical and Antinutritional Properties of Black Turtle Bean (Phaseolus Vulgaris L.) Seeds FlourUgoStanNo ratings yet
- Cosmetics: Coffee Silverskin: A Review On Potential Cosmetic ApplicationsDocument11 pagesCosmetics: Coffee Silverskin: A Review On Potential Cosmetic ApplicationsDiego Enrique Flores ValdiviaNo ratings yet
- Supercritical Antisolvent Fractionation of Lignans From The EthanolDocument7 pagesSupercritical Antisolvent Fractionation of Lignans From The Ethanolantonio troiloNo ratings yet
- Waxing As A Method of Preservation of Plantain (Musa Physicochemical Properties and Amylose Content of Plantain StarchDocument9 pagesWaxing As A Method of Preservation of Plantain (Musa Physicochemical Properties and Amylose Content of Plantain StarchgabriellexNo ratings yet
- Bioresource Technology: Iwona Gajda, John Greenman, Chris Melhuish, Carlo Santoro, Ioannis IeropoulosDocument5 pagesBioresource Technology: Iwona Gajda, John Greenman, Chris Melhuish, Carlo Santoro, Ioannis IeropoulosindahNo ratings yet
- Manfaat AntosianinDocument6 pagesManfaat AntosianinindahNo ratings yet
- Pelarut Ekstrak AntosianinDocument5 pagesPelarut Ekstrak AntosianinindahNo ratings yet
- Interaction of Carbohydrates With Alcohol Dehydrogenase: Effect On Enzyme ActivityDocument5 pagesInteraction of Carbohydrates With Alcohol Dehydrogenase: Effect On Enzyme ActivityindahNo ratings yet
- 1 s2.0 S0896844615300218 MainDocument8 pages1 s2.0 S0896844615300218 MainindahNo ratings yet
- Journal of Molecular StructureDocument7 pagesJournal of Molecular StructureindahNo ratings yet
- Week 2 Practicum ReflectionDocument2 pagesWeek 2 Practicum Reflectionapi-287584590No ratings yet
- Pulse and Digital Circuits by YoganarasimhanDocument4 pagesPulse and Digital Circuits by YoganarasimhanVignesh0% (2)
- Importance of Classroom ManagementDocument3 pagesImportance of Classroom Managementluzza2No ratings yet
- Office Performance Commitment and Review (OPCRF) : Department of EducationDocument3 pagesOffice Performance Commitment and Review (OPCRF) : Department of EducationHero LaguitNo ratings yet
- Unit-4-Methods of TrainingDocument40 pagesUnit-4-Methods of TrainingManoj KumarNo ratings yet
- Enable TDE in Oracle GG Classic ModeDocument5 pagesEnable TDE in Oracle GG Classic ModeAvinash SinghNo ratings yet
- Intertek Larson AlucoilDocument13 pagesIntertek Larson AlucoilzsmithNo ratings yet
- System Setup Ex SeriesDocument322 pagesSystem Setup Ex SeriesWaldon HendricksNo ratings yet
- Designing Organizational Structure (Adaptive Designs)Document27 pagesDesigning Organizational Structure (Adaptive Designs)haseeb0% (1)
- Gin Pole MANUALDocument13 pagesGin Pole MANUALHarish BhattNo ratings yet
- TsinghuaDocument46 pagesTsinghuabrendan lanzaNo ratings yet
- Tn-Fee Structure 2022 - 28.05.2022Document1 pageTn-Fee Structure 2022 - 28.05.2022MSPET TrustNo ratings yet
- Part I Pre-BoardDocument11 pagesPart I Pre-Boardjo-an gidoNo ratings yet
- Operator OverloadingDocument6 pagesOperator OverloadingkainatNo ratings yet
- Nacsport Scout Plus Manual enDocument131 pagesNacsport Scout Plus Manual enKoall Tchang LindienNo ratings yet
- PELSDenverSlides PDFDocument71 pagesPELSDenverSlides PDFFernando Sobrino-Manzanares MasNo ratings yet
- Dural Venous Sinus Azam SirDocument6 pagesDural Venous Sinus Azam SirLaserdotNo ratings yet
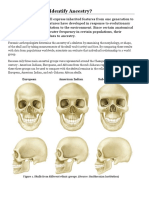
- Activity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanDocument5 pagesActivity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanpoovenmoonoosamyNo ratings yet
- Operational Guidelines On School Health Programme Under Ayushman Bharat PDFDocument40 pagesOperational Guidelines On School Health Programme Under Ayushman Bharat PDFJones PhoneNo ratings yet
- Deep Gas Well Completion Practices: H. E. Lindsey, JR., Member AIME, MWL Tool and Supply CoDocument13 pagesDeep Gas Well Completion Practices: H. E. Lindsey, JR., Member AIME, MWL Tool and Supply Cosabilco13No ratings yet
- Recipe Book: Classic Tinolang Manok RecipeDocument6 pagesRecipe Book: Classic Tinolang Manok RecipeCatherine MangloNo ratings yet
- Tekin Iftar2017 PDFDocument21 pagesTekin Iftar2017 PDFVictor Martin CobosNo ratings yet
- Topical Corticosteroid-Induced Rosacea-Like Dermatitis: A Clinical Study of 110 CasesDocument5 pagesTopical Corticosteroid-Induced Rosacea-Like Dermatitis: A Clinical Study of 110 CasesD'nesh ReYyNo ratings yet
- Paringit v. Global Gateway DigestDocument4 pagesParingit v. Global Gateway DigestBong OrduñaNo ratings yet
- Toyota Forklift 1kd Ce302 Diesel Engine Repair ManualDocument22 pagesToyota Forklift 1kd Ce302 Diesel Engine Repair Manualtamimaldonado110395eno100% (65)
- Coastal Nursing Tourism To Improve Tourist Safety in Tourist AttractionDocument10 pagesCoastal Nursing Tourism To Improve Tourist Safety in Tourist AttractionihwanNo ratings yet
- Indoor Unit/Outdoor Unit: Split Type Air ConditionerDocument23 pagesIndoor Unit/Outdoor Unit: Split Type Air ConditionerMuhamad PriyatnaNo ratings yet