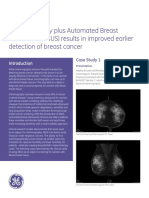
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- TitaniumDocument1 pageTitaniumErwinKSjahAlamNo ratings yet
- Size of UniverseDocument1 pageSize of UniverseErwinKSjahAlamNo ratings yet
- Size of UniverseDocument1 pageSize of UniverseErwinKSjahAlamNo ratings yet
- Asymptomatic Primary Hyper Parathyroid IsmDocument6 pagesAsymptomatic Primary Hyper Parathyroid IsmErwinKSjahAlamNo ratings yet
- English Training For Nurses and Hospital StaffDocument73 pagesEnglish Training For Nurses and Hospital Staffviaerea100% (1)
- Research Paper About ObesityDocument9 pagesResearch Paper About ObesityProsperoProllamante100% (1)
- Benign Skin LesionsDocument17 pagesBenign Skin Lesionsgarfield1No ratings yet
- ETT Seminar - Isotopes in MedicineDocument71 pagesETT Seminar - Isotopes in MedicineisocenterNo ratings yet
- Pediatrics CLIPP From QuizletDocument9 pagesPediatrics CLIPP From QuizletJoe ConeNo ratings yet
- Hepatitis B: Pathophysiology and TransmissionDocument6 pagesHepatitis B: Pathophysiology and TransmissionEndrianus Jaya PutraNo ratings yet
- Model Set MCQDocument25 pagesModel Set MCQAnugat JenaNo ratings yet
- Medical Imaging WebquestDocument8 pagesMedical Imaging Webquestapi-262193618No ratings yet
- Bethesda System For ThyroidDocument61 pagesBethesda System For ThyroidAkanksha KushwahNo ratings yet
- AllMCQsSurg2021 1Document378 pagesAllMCQsSurg2021 1AkanshaNo ratings yet
- NCM 102 Final ExamsDocument27 pagesNCM 102 Final ExamsAnonymous DZ8ZPz5100% (2)
- Bone Marrow Results 1Document7 pagesBone Marrow Results 1Top MusicNo ratings yet
- Spiritual Care in Hospice and Palliative CareDocument6 pagesSpiritual Care in Hospice and Palliative CareKrisnaNo ratings yet
- Global Saini Tomayo Case Study FinalDocument4 pagesGlobal Saini Tomayo Case Study FinalAsim AliNo ratings yet
- Sveučilište U Mostaru (Napisi Puno Ime Fakulteta I Smjer Ako To Nesto Ima)Document7 pagesSveučilište U Mostaru (Napisi Puno Ime Fakulteta I Smjer Ako To Nesto Ima)Lucija PavkovicNo ratings yet
- Tanzania STG 052013-Copy 1544379670122Document220 pagesTanzania STG 052013-Copy 1544379670122Waqar WikiNo ratings yet
- Summerassignment2 SurvivalofsickestDocument3 pagesSummerassignment2 Survivalofsickestapi-196665276No ratings yet
- Chapter 1: An Overview of Clinical LaboratoryDocument13 pagesChapter 1: An Overview of Clinical LaboratoryAshley Tañamor100% (2)
- Stroke - Final ReportDocument16 pagesStroke - Final ReportgolokipokNo ratings yet
- SBI CSR Projects - BAM18040 - ARUN DASDocument53 pagesSBI CSR Projects - BAM18040 - ARUN DASParthadeep SharmaNo ratings yet
- Screening IndustryDocument2 pagesScreening Industryrahaman.maNo ratings yet
- Rituximab MonographDocument11 pagesRituximab MonographIrma Nurtiana SyafitriNo ratings yet
- AKEEKADocument8 pagesAKEEKAJabbar AbdulNo ratings yet
- Protocol Standing Order TBDocument33 pagesProtocol Standing Order TByuniNo ratings yet
- Tongue &homoeopathic Remedies PDFDocument2 pagesTongue &homoeopathic Remedies PDFSheikh HabibiNo ratings yet
- 7 08 Ristic RDocument13 pages7 08 Ristic RsucrooNo ratings yet
- Water Excess SyndromeDocument5 pagesWater Excess SyndromeQueenie Velasquez Reinoso JacksonNo ratings yet
- Tumours and Inflammatory Lesions of The Anal Dawson2015Document12 pagesTumours and Inflammatory Lesions of The Anal Dawson2015DannyNo ratings yet
- Congenital Heart DseDocument14 pagesCongenital Heart DseJedzRNNo ratings yet
- Surgery 2 Case ReportDocument12 pagesSurgery 2 Case ReportElvis NgNo ratings yet