You might also like
- Physiology and BiochemistryFrom EverandPhysiology and BiochemistryGeoffrey BourneNo ratings yet
- Nursing-AnaPhy-Muscular System Upper BodyDocument21 pagesNursing-AnaPhy-Muscular System Upper BodyGail Chantel Spring PerlasNo ratings yet
- Tendon Neuroplastic TrainingDocument9 pagesTendon Neuroplastic TrainingLara VitorNo ratings yet
- AIS and YogaDocument3 pagesAIS and Yogasale18No ratings yet
- Eccentric Vs Concentric Training Muscle GrowthDocument8 pagesEccentric Vs Concentric Training Muscle GrowthAres Santiago R. NoguedaNo ratings yet
- Chondromalacia Patella RehabDocument1 pageChondromalacia Patella Rehabjntownsend3404No ratings yet
- Ankle Rehabilitation: DR - Zahoor Ahmad Assistant Professor NCS UniversityDocument25 pagesAnkle Rehabilitation: DR - Zahoor Ahmad Assistant Professor NCS UniversityZahoor AhmadNo ratings yet
- Groin Strain Rehabilitation PDFDocument3 pagesGroin Strain Rehabilitation PDFOscar NgNo ratings yet
- Asana Based Exercises Low Back PainDocument14 pagesAsana Based Exercises Low Back PainAutumn Carter100% (1)
- EMG in Weightlifting Performance During The SnatchDocument5 pagesEMG in Weightlifting Performance During The SnatchsafaaismaeelNo ratings yet
- Massage and Performance Recovery A Meta-Analytical Review Sports MedicineDocument22 pagesMassage and Performance Recovery A Meta-Analytical Review Sports MedicineBelén María Vega AladueñaNo ratings yet
- Strapping TechniquesDocument19 pagesStrapping Techniqueswan100% (1)
- Effects of Petrissage Massage On Fatigue and ExercDocument8 pagesEffects of Petrissage Massage On Fatigue and ExercJGRBBKDVRNo ratings yet
- Self Myofascial ReleaseDocument22 pagesSelf Myofascial ReleaseMuhammad Fahmy100% (1)
- 4 PTY121 Exercise PrescriptionDocument56 pages4 PTY121 Exercise PrescriptionsenoNo ratings yet
- Bone Fracture & RemodellingDocument39 pagesBone Fracture & Remodellingokware rodrickNo ratings yet
- Stretching Exercise Therapy and Primary Dysmenorrhea - Nursing PerspectivesDocument4 pagesStretching Exercise Therapy and Primary Dysmenorrhea - Nursing PerspectivesIOSRjournalNo ratings yet
- Core Stability Rippetoe PDFDocument4 pagesCore Stability Rippetoe PDFSylvia GraceNo ratings yet
- Multidisciplinary Sports Injury Recovery ProcessDocument5 pagesMultidisciplinary Sports Injury Recovery ProcessHoorab SiddiqaNo ratings yet
- Met JGDocument3 pagesMet JGthoracic555No ratings yet
- Kinetic Chain Exercises - Open Versus ClosedDocument4 pagesKinetic Chain Exercises - Open Versus Closedamirreza jmNo ratings yet
- Notes For PTA NPTEFFDocument19 pagesNotes For PTA NPTEFFFarhana Rashena100% (1)
- Common Dance InjuriesDocument5 pagesCommon Dance InjuriesJeyceeNo ratings yet
- Principles of Marathon TrainingDocument6 pagesPrinciples of Marathon TrainingaloordominicNo ratings yet
- Pilates Exercises (Inglés) (Artículo) - Salvi ShahDocument8 pagesPilates Exercises (Inglés) (Artículo) - Salvi ShahJose Luis Ramos MolloNo ratings yet
- Active Recovery PlanDocument4 pagesActive Recovery PlanIwatobiNo ratings yet
- A Foot Core Approach To Treating Plantar Fasciitis SmallDocument4 pagesA Foot Core Approach To Treating Plantar Fasciitis SmallLeticia MeleganNo ratings yet
- Apostila de Tantsu CradleDocument10 pagesApostila de Tantsu CradlereikianosNo ratings yet
- Circuit Training HandoutsDocument4 pagesCircuit Training HandoutsGwyneth Bunda100% (1)
- Hip Assessment Worksheet Page2Document1 pageHip Assessment Worksheet Page2David GoodwinNo ratings yet
- Strengthening Exercises For Calf Muscles: Steps To Strengthen The Gastrocnemius and Soleus (Calf) MusclesDocument2 pagesStrengthening Exercises For Calf Muscles: Steps To Strengthen The Gastrocnemius and Soleus (Calf) MusclesChristopher Ozil McanuffNo ratings yet
- Muscle CrampsDocument7 pagesMuscle Crampsapi-439746419No ratings yet
- Intro To OMT: Stacia Sloane, DODocument37 pagesIntro To OMT: Stacia Sloane, DOJeremy100% (1)
- Efficacy of Neck Stabilization Exercises For Neck Pain: A Randomized Controlled StudyDocument6 pagesEfficacy of Neck Stabilization Exercises For Neck Pain: A Randomized Controlled StudyAnonymous mWAaDZmlJwNo ratings yet
- WEEK 2 STRESS BREAKDocument6 pagesWEEK 2 STRESS BREAKHey itsJamNo ratings yet
- A Technical Description of The Pull in WeightliftingDocument7 pagesA Technical Description of The Pull in WeightliftingpluckypuckNo ratings yet
- Theory For MassageDocument16 pagesTheory For MassageAnnaya KhanNo ratings yet
- Biomechanics of The Shoulder ComplexDocument83 pagesBiomechanics of The Shoulder ComplexpiciliviNo ratings yet
- Assessment of KneeDocument113 pagesAssessment of KneeIqra Iftikhar100% (1)
- WEEK 6 Muscular SystemDocument17 pagesWEEK 6 Muscular SystemMarjorie Lyka LingatNo ratings yet
- StretchesDocument44 pagesStretchesdoktorgreenNo ratings yet
- ACL Rehab Guidelines for Early Range of Motion and StrengtheningDocument4 pagesACL Rehab Guidelines for Early Range of Motion and Strengtheningfia0711No ratings yet
- Functional Exercise ProgressionDocument6 pagesFunctional Exercise ProgressionAnastasia Skamagki100% (1)
- Lecture No. 1 Vertebral Motion DynamicsDocument67 pagesLecture No. 1 Vertebral Motion DynamicsSYED ALI HUSSAIN100% (1)
- Relax Massage - TranslatedDocument30 pagesRelax Massage - Translatedsoula90No ratings yet
- KneeDocument2 pagesKneedrkmsk100% (1)
- Overhead Throwing: Biomechanics and PathologyDocument7 pagesOverhead Throwing: Biomechanics and Pathologyrapannika100% (1)
- Stretch PDFDocument14 pagesStretch PDFCarolina B. SánchezNo ratings yet
- Flexibility Training: Len Kravitz, Ph.D. and Vivian H. Heyward, PH.DDocument12 pagesFlexibility Training: Len Kravitz, Ph.D. and Vivian H. Heyward, PH.Dgschiro100% (1)
- Mobility and Joint Function: 6 Key ExercisesDocument22 pagesMobility and Joint Function: 6 Key ExercisesGhost GamerxNo ratings yet
- Current Concepts of Plyometric ExerciseDocument27 pagesCurrent Concepts of Plyometric ExerciseMustafid Alwi AlvaroNo ratings yet
- Exercise 2 Activity 3 Muscle SummationDocument13 pagesExercise 2 Activity 3 Muscle SummationElvira BasilioNo ratings yet
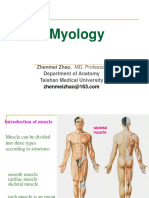
- Myology: Zhenmei Zhao, MD, Professor Department of Anatomy Taishan Medical UniversityDocument123 pagesMyology: Zhenmei Zhao, MD, Professor Department of Anatomy Taishan Medical UniversityAkshit SetiaNo ratings yet
- Scapular Winging With Rehab-SportsMedDocument7 pagesScapular Winging With Rehab-SportsMedWasemBhatNo ratings yet
- Meniscus Tears - Cartilage Damage: KneeDocument9 pagesMeniscus Tears - Cartilage Damage: KneedheenadayalanNo ratings yet
- Biomechanics of YogaDocument45 pagesBiomechanics of YogaKrishnakumar PNo ratings yet
- Frozen ShoulderDocument10 pagesFrozen ShoulderqimindraNo ratings yet
- Pilates InserviceDocument11 pagesPilates InserviceToni MansfieldNo ratings yet
- General RelaxationDocument4 pagesGeneral RelaxationMANANNo ratings yet
- Cervical Artery Dysfunction: Implications For Physiotherapy Diagnosis and ManagementDocument260 pagesCervical Artery Dysfunction: Implications For Physiotherapy Diagnosis and ManagementrapannikaNo ratings yet
- Cervical Artery Dysfunction and ManipulationDocument8 pagesCervical Artery Dysfunction and ManipulationrapannikaNo ratings yet
- De Las Penas V17N3Document9 pagesDe Las Penas V17N3rapannikaNo ratings yet
- Clinical Examination of Patients With Headache ComplaintsDocument157 pagesClinical Examination of Patients With Headache ComplaintsrapannikaNo ratings yet
- V17N4 AnloagueDocument8 pagesV17N4 AnloaguerapannikaNo ratings yet
- Low Back Pain Presentation IMPDocument114 pagesLow Back Pain Presentation IMPrapannika100% (3)
- Editorial: A New Research Model For Orthopaedic Manual Physical TherapyDocument3 pagesEditorial: A New Research Model For Orthopaedic Manual Physical Therapyrapannika100% (2)
- Lumbopelvic Stability: SyllabusDocument13 pagesLumbopelvic Stability: Syllabusrapannika100% (3)
- Cervicogenic Dizziness and DDXDocument64 pagesCervicogenic Dizziness and DDXrapannika100% (3)
- Editorial: Thurst Manipulation in United States Entry-Level Physical Therapy Curricular ContentDocument3 pagesEditorial: Thurst Manipulation in United States Entry-Level Physical Therapy Curricular Contentrapannika100% (2)
- Cervical Artery PresentationDocument198 pagesCervical Artery Presentationrapannika100% (3)
- Presentation On Advances in The Physical Therapy Diagnosis, Prognosis, and Management of Patients With Low Back PainDocument85 pagesPresentation On Advances in The Physical Therapy Diagnosis, Prognosis, and Management of Patients With Low Back Painrapannika100% (2)
- Adding Thrust Manipulation To Entry-Level Physical Therapy Curricula: EditorialDocument4 pagesAdding Thrust Manipulation To Entry-Level Physical Therapy Curricula: Editorialrapannika100% (2)
- Sacroiliac Joint SyllabusDocument16 pagesSacroiliac Joint Syllabusrapannika100% (2)
- Editorial: Clinical Prediction Rules in Manual Physical TherapyDocument4 pagesEditorial: Clinical Prediction Rules in Manual Physical Therapyrapannika100% (2)
- Editorial: Manual Therapy in ChildrenDocument3 pagesEditorial: Manual Therapy in Childrenrapannika100% (3)
- Editorial: Physical Therapy and Chiropractic and The Right To ManipulateDocument12 pagesEditorial: Physical Therapy and Chiropractic and The Right To Manipulaterapannika100% (2)
- Lumbopelvic Monograph: Pathology, Examination, Diagnosis, and ManagementDocument47 pagesLumbopelvic Monograph: Pathology, Examination, Diagnosis, and Managementrapannika100% (2)
- Guest Editorial Evidence-Based Physical Therapy PracticeDocument1 pageGuest Editorial Evidence-Based Physical Therapy Practicerapannika100% (2)
- Letter To The Editor - Substance AbuseDocument2 pagesLetter To The Editor - Substance Abuserapannika100% (2)
- Editorial: Chiropractic Subluxation and Implications For Manual Physical TherapyDocument3 pagesEditorial: Chiropractic Subluxation and Implications For Manual Physical Therapyrapannika100% (3)
- Sacroiliac Joint Dysfunction: A Narrative Review of Evidence-Based DiagnosisDocument19 pagesSacroiliac Joint Dysfunction: A Narrative Review of Evidence-Based Diagnosisrapannika100% (5)
- Editorial Evidence-Based Manual Physical TherapyDocument2 pagesEditorial Evidence-Based Manual Physical Therapyrapannika100% (2)
- Coupled Motions of The Lumbar Spine: A Narrative ReviewDocument5 pagesCoupled Motions of The Lumbar Spine: A Narrative Reviewrapannika100% (2)
- Ergogenic Substances: A MonographDocument40 pagesErgogenic Substances: A Monographrapannika100% (2)
- Lumbopelvic Monograph: Anatomy and BiomechanicsDocument46 pagesLumbopelvic Monograph: Anatomy and Biomechanicsrapannika100% (2)
- Tendon Injury: A Narrative ReviewDocument10 pagesTendon Injury: A Narrative Reviewrapannika100% (2)
- Theravital Bicycle Trainer: Quasi-Experimental Trail in A Geriatric PopulationDocument6 pagesTheravital Bicycle Trainer: Quasi-Experimental Trail in A Geriatric Populationrapannika100% (2)
- Scaphoid Fracture: A Case ReportDocument5 pagesScaphoid Fracture: A Case Reportrapannika100% (2)
- Tendon Injury: A Narrative ReviewDocument10 pagesTendon Injury: A Narrative Reviewrapannika100% (2)
- Questions 1 Through 10 Are Based On The Following PassageDocument5 pagesQuestions 1 Through 10 Are Based On The Following PassageSarjitoNo ratings yet
- Biolis24i CatalogueDocument8 pagesBiolis24i CataloguekadirucaNo ratings yet
- Acitretin ACH and MDSDocument5 pagesAcitretin ACH and MDSWei Sheng ChongNo ratings yet
- Uhs MCQS Globular ProteinsDocument16 pagesUhs MCQS Globular ProteinsRaza AliNo ratings yet
- Grade 8 Science: Learning Area: Living Things and Their Environment Quarter: Fourth QuarterDocument4 pagesGrade 8 Science: Learning Area: Living Things and Their Environment Quarter: Fourth QuarterLexter CarmenNo ratings yet
- High School Biology ReviewerDocument26 pagesHigh School Biology ReviewerPatricia SioNo ratings yet
- WWW Mcqsurgery ComDocument4 pagesWWW Mcqsurgery ComSajag GuptaNo ratings yet
- Observe Science Magazine 1st EditionDocument31 pagesObserve Science Magazine 1st Editionapi-408725114No ratings yet
- YogaDocument12 pagesYogaRahfyahamedNo ratings yet
- Assignment No. 1Document7 pagesAssignment No. 1heidisjdhdhdhNo ratings yet
- Chitosan Nanoparticles PHD ThesisDocument6 pagesChitosan Nanoparticles PHD Thesisfjgmmmew100% (2)
- Paramolar - A Supernumerary Molar A Case Report and An OverviewDocument7 pagesParamolar - A Supernumerary Molar A Case Report and An OverviewGurudutt NayakNo ratings yet
- History Taking in OrthopedicsDocument28 pagesHistory Taking in OrthopedicsKenneth SuliyantoNo ratings yet
- University of Helsinki Dog StudyDocument10 pagesUniversity of Helsinki Dog StudySarah LightNo ratings yet
- The Intersection Between Forensic Anthropometry and Cellular Physiology For Investigations: Prospects in Low and Middle-Income CountriesDocument13 pagesThe Intersection Between Forensic Anthropometry and Cellular Physiology For Investigations: Prospects in Low and Middle-Income CountriesIJAR JOURNALNo ratings yet
- HLTH 1020 Dietary Analysis Final AssignmentDocument11 pagesHLTH 1020 Dietary Analysis Final Assignmentapi-438184229No ratings yet
- Extended Response 6 QDocument2 pagesExtended Response 6 Q12-057 MOHAMMAD MUSHFIQUR RAHMANNo ratings yet
- Pancreatic Cancer: A Review of Risk Factors, Diagnosis, and TreatmentDocument13 pagesPancreatic Cancer: A Review of Risk Factors, Diagnosis, and TreatmentNoy FaridaNo ratings yet
- Biology NotesDocument102 pagesBiology Notesajabgul123493No ratings yet
- Clinical Pharmacology of Caffeine Citrate in PreteDocument8 pagesClinical Pharmacology of Caffeine Citrate in PreteVictoria SolomonNo ratings yet
- Abortions Dohbit Yaounde 2007 PDFDocument26 pagesAbortions Dohbit Yaounde 2007 PDFBadlina Fitrianisa YulianingrumNo ratings yet
- CELLS For Class8Document15 pagesCELLS For Class8cbseiscNo ratings yet
- Retraso Global Del DesarrolloDocument18 pagesRetraso Global Del Desarrolloenypaola19No ratings yet
- C - The Microwave Syndrome - 13 CategoriesDocument4 pagesC - The Microwave Syndrome - 13 CategoriesEmf RefugeeNo ratings yet
- Recent Advances in Retinoblastoma Management and GeneticsDocument12 pagesRecent Advances in Retinoblastoma Management and GeneticsDr Pranesh BalasubramaniamNo ratings yet
- Associations of Adiposity, Circulating Protein Biomarkers, and Risk of Major Vascular DiseasesDocument11 pagesAssociations of Adiposity, Circulating Protein Biomarkers, and Risk of Major Vascular DiseasesYuzhuo WangNo ratings yet
- Integrating Genomics Into Healthcare: A Global ResponsibilityDocument14 pagesIntegrating Genomics Into Healthcare: A Global ResponsibilityMattNo ratings yet
- Somatoform DisordersDocument43 pagesSomatoform DisordersChristopher El MouhayyarNo ratings yet
- AmniocentesisDocument15 pagesAmniocentesisGYPSY CAT100% (1)
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument15 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsArya ChowdhuryNo ratings yet