You might also like
- Observation of Live Chromatin Dynamics in Cells Via 3D Localization Microscopy Using Tetrapod Point Spread FunctionsDocument14 pagesObservation of Live Chromatin Dynamics in Cells Via 3D Localization Microscopy Using Tetrapod Point Spread FunctionsnimasolhjooNo ratings yet
- TMP 34 FDDocument17 pagesTMP 34 FDFrontiersNo ratings yet
- Azucena 2010Document12 pagesAzucena 2010mahmoud khedrNo ratings yet
- Measuring Easily Electron Plasma Densities in Gases Produced by Ultrashort Lasers and FilamentsDocument6 pagesMeasuring Easily Electron Plasma Densities in Gases Produced by Ultrashort Lasers and FilamentsΤάσος ΚουλουκλίδηςNo ratings yet
- Dynamic Three-Dimensional Tracking of Single Fluorescent Nanoparticles Deep Inside Living TissueDocument11 pagesDynamic Three-Dimensional Tracking of Single Fluorescent Nanoparticles Deep Inside Living TissueRicardo AlvaradoNo ratings yet
- Oe 16 009884Document11 pagesOe 16 009884benetorio.tNo ratings yet
- Boe 7 9 3355Document22 pagesBoe 7 9 3355api-310329774No ratings yet
- 2007 - Heifetz Et Al. - Subdiffraction Optical Resolution of A Gold Nanosphere Located Within The Nanojet of A Mie-Resonant Dielectric MDocument9 pages2007 - Heifetz Et Al. - Subdiffraction Optical Resolution of A Gold Nanosphere Located Within The Nanojet of A Mie-Resonant Dielectric MClaudio BiaginiNo ratings yet
- Biophoton Papers PDFDocument5 pagesBiophoton Papers PDFBrightchip TechnologiesNo ratings yet
- TMP 1 F64Document18 pagesTMP 1 F64FrontiersNo ratings yet
- Handbook of Biomedical Nonlinear Optical Microscopy 2008 EbookDocument895 pagesHandbook of Biomedical Nonlinear Optical Microscopy 2008 EbookPriscilla González MatamorosNo ratings yet
- Applications in Clinical and Biomedical Research. Technical University of DenmarkDocument2 pagesApplications in Clinical and Biomedical Research. Technical University of DenmarkMIRANDARISKINo ratings yet
- Millisecond Infrared Laser Pulses Depolarize and Elicit Action Potentials On In-Vitro Dorsal Root Ganglion NeuronsDocument11 pagesMillisecond Infrared Laser Pulses Depolarize and Elicit Action Potentials On In-Vitro Dorsal Root Ganglion Neuronsender000No ratings yet
- Oe 13 23 9361Document13 pagesOe 13 23 9361Aggelos StefopoulosNo ratings yet
- Fluorescence Microscopy: Super-Resolution and other Novel TechniquesFrom EverandFluorescence Microscopy: Super-Resolution and other Novel TechniquesAnda CorneaNo ratings yet
- Speculations About Bystander and BiophotonsDocument3 pagesSpeculations About Bystander and BiophotonsGabrielNo ratings yet
- Imaging Cone Photoreceptors in Three Dimensions and in Time Using Ultrahigh Resolution Optical Coherence Tomography With Adaptive OpticsDocument16 pagesImaging Cone Photoreceptors in Three Dimensions and in Time Using Ultrahigh Resolution Optical Coherence Tomography With Adaptive OpticsChuanlin FanNo ratings yet
- Biophoton - WikipediaDocument6 pagesBiophoton - WikipediaCarl MacCordNo ratings yet
- Mueller Decomposition Images For Cervical Tissue: Potential For Discriminating Normal and Dysplastic StatesDocument10 pagesMueller Decomposition Images For Cervical Tissue: Potential For Discriminating Normal and Dysplastic StatesSiddaraj UppinNo ratings yet
- Field-Portable Reflection and Transmission Microscopy Based On Lensless HolographyDocument10 pagesField-Portable Reflection and Transmission Microscopy Based On Lensless HolographyShiva GaddamNo ratings yet
- Paper 18researchDocument15 pagesPaper 18researchjeremybregmanNo ratings yet
- Integration of Optical Components On-Chip For Scattering and Fluorescence Detection in An Optofluidic DeviceDocument10 pagesIntegration of Optical Components On-Chip For Scattering and Fluorescence Detection in An Optofluidic DeviceDaniel MăriuțaNo ratings yet
- 2012 BOE v3 n8 p1774 LiewDocument13 pages2012 BOE v3 n8 p1774 LiewYih Miin LiewNo ratings yet
- 論文02Document8 pages論文02張贇No ratings yet
- Speculations About Bystander and Biophotons - PMCDocument4 pagesSpeculations About Bystander and Biophotons - PMCCarl MacCordNo ratings yet
- Ex-Vivo Characterization of Human Colon Cancer: by Mueller Polarimetric ImagingDocument12 pagesEx-Vivo Characterization of Human Colon Cancer: by Mueller Polarimetric ImagingnuralitarpNo ratings yet
- Oe 12 14 3055 PDFDocument11 pagesOe 12 14 3055 PDFFei YiNo ratings yet
- Advances in Microscopy Techniques: Review ArticleDocument9 pagesAdvances in Microscopy Techniques: Review Articlekajol22fulNo ratings yet
- Determining The Optical Properties of Gelatine GelDocument17 pagesDetermining The Optical Properties of Gelatine GelCorynaNo ratings yet
- Chaos in Ocular Aberration Dynamics of The Human Eye: Karen M. Hampson and Edward A. H. MallenDocument15 pagesChaos in Ocular Aberration Dynamics of The Human Eye: Karen M. Hampson and Edward A. H. MallenFrank smith Condor loarteNo ratings yet
- Glass Rev2Document19 pagesGlass Rev2lobitos.1188No ratings yet
- On-Chip Generation of Time-And Wavelength-Division Multiplexed Multiple Time-Bin EntanglementDocument10 pagesOn-Chip Generation of Time-And Wavelength-Division Multiplexed Multiple Time-Bin EntanglementBukhariNo ratings yet
- Three-Dimensional Optical Memory Using A Human FingernailDocument8 pagesThree-Dimensional Optical Memory Using A Human FingernailPuja srinija AtukuriNo ratings yet
- Optik 68Document15 pagesOptik 68z.umul9031No ratings yet
- Magnetic Resonance Imaging, Microscopy, and Spectroscopy of The Central Nervous System in Experimental AnimalsDocument15 pagesMagnetic Resonance Imaging, Microscopy, and Spectroscopy of The Central Nervous System in Experimental Animalsxmd2011No ratings yet
- Label-Free Imaging of Membrane Potential Using Membrane ElectromotilityDocument8 pagesLabel-Free Imaging of Membrane Potential Using Membrane ElectromotilityNur AnindyaNo ratings yet
- Publication List 200912 f2Document5 pagesPublication List 200912 f2MBPL webNo ratings yet
- Kabashin 2009Document14 pagesKabashin 2009Caio VianaNo ratings yet
- Tissue Optics and Photonics: Biological Tissue StructuresDocument19 pagesTissue Optics and Photonics: Biological Tissue StructuresAnonymous efLc7hSit5No ratings yet
- Multi-Band Metamaterial Absorber Based On The Arrangement of Donut-Type ResonatorsDocument12 pagesMulti-Band Metamaterial Absorber Based On The Arrangement of Donut-Type ResonatorsanhtrinhndNo ratings yet
- Ultrasound: Its Applications in Medicine and BiologyFrom EverandUltrasound: Its Applications in Medicine and BiologyNo ratings yet
- Thesis Haichun Configuratia Electronica TabelDocument229 pagesThesis Haichun Configuratia Electronica TabelAngela StefanNo ratings yet
- Microbeam Analysis in BiologyFrom EverandMicrobeam Analysis in BiologyClaude LecheneNo ratings yet
- Oe 21 3 3213Document12 pagesOe 21 3 3213Adarsh VishwakarmaNo ratings yet
- Three Dimensional Microanatomy of Cells and Tissue Surfaces: Proceedings of the Symposium on Three Dimensional Microanatomy held in Mexico City, Mexico, August 17-23, 1980From EverandThree Dimensional Microanatomy of Cells and Tissue Surfaces: Proceedings of the Symposium on Three Dimensional Microanatomy held in Mexico City, Mexico, August 17-23, 1980Delmas J. AllenRating: 5 out of 5 stars5/5 (1)
- Spectroscopy in Biology and Chemistry: Neutron, X-Ray, LaserFrom EverandSpectroscopy in Biology and Chemistry: Neutron, X-Ray, LaserSow-Hsin ChenNo ratings yet
- Microphotography - A ReviewDocument3 pagesMicrophotography - A ReviewIJAR JOURNALNo ratings yet
- The Study of Electrokinetic Properties of Buccal EpitheliumDocument5 pagesThe Study of Electrokinetic Properties of Buccal Epitheliumgeorgbell5No ratings yet
- Single-Arm Three-Wave Interferometer For Measuring Dispersion of Short Lengths of FiberDocument13 pagesSingle-Arm Three-Wave Interferometer For Measuring Dispersion of Short Lengths of Fiberyuvabala2002123No ratings yet
- Quantifying tissue mechanical properties using photoplethysmographyDocument14 pagesQuantifying tissue mechanical properties using photoplethysmographyCalm YourselfNo ratings yet
- Creath-Schwartz - Biophoton Interaction in Biological SystemsDocument10 pagesCreath-Schwartz - Biophoton Interaction in Biological SystemsGabrielNo ratings yet
- Micro-And Nano-CT For The Study of Bone Ultrastructure: Imaging (T Lang and F Wehrli, Section Editors)Document10 pagesMicro-And Nano-CT For The Study of Bone Ultrastructure: Imaging (T Lang and F Wehrli, Section Editors)yhlnjpcjNo ratings yet
- Membrane Potential Imaging in The Nervous System and Heart: Marco Canepari Dejan Zecevic Olivier Bernus EditorsDocument511 pagesMembrane Potential Imaging in The Nervous System and Heart: Marco Canepari Dejan Zecevic Olivier Bernus EditorsHORACIO VARGASNo ratings yet
- Si-CMOS Compatible Materials and Devices For mid-IR MicrophotonicsDocument14 pagesSi-CMOS Compatible Materials and Devices For mid-IR MicrophotonicsMegan MakelaNo ratings yet
- Lima Boe p1069 2011 Is To OctDocument13 pagesLima Boe p1069 2011 Is To OctIvan LimaNo ratings yet
- BOE Puszka CorrectedDocument15 pagesBOE Puszka CorrectedEsraa AymanNo ratings yet
- The Study of Electrokinetic Properties of Buccal EpitheliumDocument4 pagesThe Study of Electrokinetic Properties of Buccal Epitheliumgeorgbell5No ratings yet
- Taller Microscopia 1Document46 pagesTaller Microscopia 1Erika PereaNo ratings yet
- UFO FILES Black Box Ufo SecretsDocument10 pagesUFO FILES Black Box Ufo SecretsNomad XNo ratings yet
- Synopsis On Training & DevelopmentDocument6 pagesSynopsis On Training & DevelopmentArchi gupta86% (14)
- Indiana University PressDocument33 pagesIndiana University Pressrenato lopesNo ratings yet
- .Design and Development of Motorized Multipurpose MachineDocument3 pages.Design and Development of Motorized Multipurpose MachineANKITA MORENo ratings yet
- Materiality of Cultural ConstructionDocument3 pagesMateriality of Cultural ConstructionPaul YeboahNo ratings yet
- Lesson 4: Mean and Variance of Discrete Random Variable: Grade 11 - Statistics & ProbabilityDocument26 pagesLesson 4: Mean and Variance of Discrete Random Variable: Grade 11 - Statistics & Probabilitynicole MenesNo ratings yet
- Chapter 2 ProblemsDocument41 pagesChapter 2 ProblemsPrakasam ArulappanNo ratings yet
- Research Article: Noise-Cancelling CMOS Active Inductor and Its Application in RF Band-Pass Filter DesignDocument8 pagesResearch Article: Noise-Cancelling CMOS Active Inductor and Its Application in RF Band-Pass Filter DesignAbhay S KochharNo ratings yet
- Government of India Sponsored Study on Small Industries Potential in Sultanpur DistrictDocument186 pagesGovernment of India Sponsored Study on Small Industries Potential in Sultanpur DistrictNisar Ahmed Ansari100% (1)
- Other Books NewDocument136 pagesOther Books NewYashRaj ChoudharyNo ratings yet
- Assignment 7Document1 pageAssignment 7sujit kcNo ratings yet
- Duobias M 200 TM Section02 Performance SpecificationDocument12 pagesDuobias M 200 TM Section02 Performance SpecificationtonytomsonNo ratings yet
- Modular Kitchen AnalysisDocument3 pagesModular Kitchen AnalysisGhanithan SubramaniamNo ratings yet
- Babylonian Mathematics (Also Known As Assyro-BabylonianDocument10 pagesBabylonian Mathematics (Also Known As Assyro-BabylonianNirmal BhowmickNo ratings yet
- Shimano 105 Gear Change ManualDocument1 pageShimano 105 Gear Change Manual1heUndertakerNo ratings yet
- Check List Valve PDFDocument2 pagesCheck List Valve PDFikan100% (1)
- Adobe Scan 25 Dec 202Document1 pageAdobe Scan 25 Dec 202Jimit ShahNo ratings yet
- Solid Solution For Catalytic Ammonia Synthesis FromDocument8 pagesSolid Solution For Catalytic Ammonia Synthesis FromAhmad AlShahrourNo ratings yet
- TransformationsRotations LessonPlanDocument6 pagesTransformationsRotations LessonPlanocsc100% (1)
- Limit of Outside Usage Outside Egypt ENDocument1 pageLimit of Outside Usage Outside Egypt ENIbrahem EmamNo ratings yet
- Green Building by Superadobe TechnologyDocument22 pagesGreen Building by Superadobe TechnologySivaramakrishnaNalluri67% (3)
- Quality OrientationDocument2 pagesQuality OrientationResearcherNo ratings yet
- Vietnamese sentence structure: key rules for SVO ordering, adjective and adverb placementDocument7 pagesVietnamese sentence structure: key rules for SVO ordering, adjective and adverb placementvickyzaoNo ratings yet
- PROPOSED ARFF BUILDINGDocument27 pagesPROPOSED ARFF BUILDINGDale Bryan S DalmacioNo ratings yet
- Property Law Supp 2016Document6 pagesProperty Law Supp 2016Tefo TshukuduNo ratings yet
- PHD Thesis - Dalong Zhao-ZnO TFT Thin Film Passivation - Noise Analysis, Hooge ParameterDocument169 pagesPHD Thesis - Dalong Zhao-ZnO TFT Thin Film Passivation - Noise Analysis, Hooge Parameterjiaxin zhangNo ratings yet
- Oisd STD 175Document59 pagesOisd STD 175yandiNo ratings yet
- Drone Hexacopter IjisrtDocument4 pagesDrone Hexacopter IjisrtDEPARTEMEN RUDALNo ratings yet
- Costallocationvideolectureslides 000XADocument12 pagesCostallocationvideolectureslides 000XAWOw Wong100% (1)
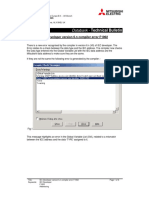
- IEC Developer version 6.n compiler error F1002Document6 pagesIEC Developer version 6.n compiler error F1002AnddyNo ratings yet