You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Slaked Lime MSDS Safety SummaryDocument7 pagesSlaked Lime MSDS Safety SummaryFurqan SiddiquiNo ratings yet
- Apc 8x Install Config Guide - rn0 - LT - enDocument162 pagesApc 8x Install Config Guide - rn0 - LT - enOney Enrique Mendez MercadoNo ratings yet
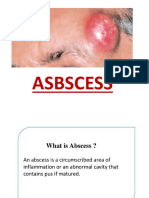
- ABSCESSDocument35 pagesABSCESSlax prajapatiNo ratings yet
- Alternate Mekton Zeta Weapon CreationDocument7 pagesAlternate Mekton Zeta Weapon CreationJavi BuenoNo ratings yet
- The Danger of Microwave TechnologyDocument16 pagesThe Danger of Microwave Technologyrey_hadesNo ratings yet
- Dip Obst (SA) Past Papers - 2020 1st Semester 1-6-2023Document1 pageDip Obst (SA) Past Papers - 2020 1st Semester 1-6-2023Neo Latoya MadunaNo ratings yet
- Introduction To Animal Science For Plant ScienceDocument63 pagesIntroduction To Animal Science For Plant ScienceJack OlanoNo ratings yet
- Synthesis, Experimental and Theoretical Characterizations of A NewDocument7 pagesSynthesis, Experimental and Theoretical Characterizations of A NewWail MadridNo ratings yet
- M-LVDT: Microminiature Displacement SensorDocument2 pagesM-LVDT: Microminiature Displacement Sensormahdi mohammadiNo ratings yet
- Arp0108 2018Document75 pagesArp0108 2018justin.kochNo ratings yet
- Cap 716 PDFDocument150 pagesCap 716 PDFjanhaviNo ratings yet
- Guide Propedevt Stomat 2c EngDocument256 pagesGuide Propedevt Stomat 2c EngJhoel Jhonatan Torres MuñozNo ratings yet
- Past The Shallows EssayDocument2 pagesPast The Shallows EssaycaitlinNo ratings yet
- Thee Correlational Study of Possittive Emotionons and Coping Strategies For Academic Stress Among CASS Studentts - updaTEDDocument23 pagesThee Correlational Study of Possittive Emotionons and Coping Strategies For Academic Stress Among CASS Studentts - updaTEDJuliet AcelNo ratings yet
- 4.5 Cargo DischargingDocument2 pages4.5 Cargo DischargingHasan Cosalev100% (1)
- General Specifications: Detail ADocument1 pageGeneral Specifications: Detail AJeniel PascualNo ratings yet
- Dr. Namrata Misra Head of Bioinnovations at KIIT UniversityDocument1 pageDr. Namrata Misra Head of Bioinnovations at KIIT Universitymanisha maniNo ratings yet
- ERS M22 PC4 FerryDocument2 pagesERS M22 PC4 FerryouakgoodNo ratings yet
- Request Baha Alert PPMPDocument3 pagesRequest Baha Alert PPMPLö Räine AñascoNo ratings yet
- Dimensional Data: For Valves and ActuatorsDocument52 pagesDimensional Data: For Valves and ActuatorsPaulNo ratings yet
- Very Easy Toeic Units 7 - 12 (Q1)Document39 pagesVery Easy Toeic Units 7 - 12 (Q1)Minh KhaiNo ratings yet
- g21 Gluta MsdsDocument3 pagesg21 Gluta Msdsiza100% (1)
- Soal Upk B Inggris PKBM WinaDocument11 pagesSoal Upk B Inggris PKBM WinaCuman MitosNo ratings yet
- Ignition System Spark Test DiagnosisDocument24 pagesIgnition System Spark Test DiagnosisMohamed l'Amine75% (4)
- Quality Control Plan Static EquipmentDocument1 pageQuality Control Plan Static EquipmentdhasdjNo ratings yet
- HTM 2025 2 (New) Ventilation in HospitalsDocument123 pagesHTM 2025 2 (New) Ventilation in HospitalsArvish RamseebaluckNo ratings yet
- WSAWLD002Document29 pagesWSAWLD002Nc BeanNo ratings yet
- The Girls Center: 2023 Workout CalendarDocument17 pagesThe Girls Center: 2023 Workout Calendark4270621No ratings yet
- Merit of RatingDocument1 pageMerit of RatingRaidasNo ratings yet
- Lease Practice QuestionsDocument4 pagesLease Practice QuestionsAbdul SamiNo ratings yet