You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Cham PowerpointDocument19 pagesCham PowerpointElizabeth GenotivaNo ratings yet
- Plaza 2019Document13 pagesPlaza 2019nicolasdizzNo ratings yet
- Consciousness: Here, There and Everywhere?: ReviewDocument18 pagesConsciousness: Here, There and Everywhere?: ReviewnicolasdizzNo ratings yet
- Neural Correlates of Consciousness - Koch 2016Document15 pagesNeural Correlates of Consciousness - Koch 2016Juan Cristobal Ruiz Diaz100% (1)
- The Neuroscientist: Brain Connectivity: Gender Makes A DifferenceDocument18 pagesThe Neuroscientist: Brain Connectivity: Gender Makes A DifferencenicolasdizzNo ratings yet
- Consciousness: Here, There and Everywhere?: ReviewDocument18 pagesConsciousness: Here, There and Everywhere?: ReviewnicolasdizzNo ratings yet
- The Neuroscientist: Brain Connectivity: Gender Makes A DifferenceDocument18 pagesThe Neuroscientist: Brain Connectivity: Gender Makes A DifferencenicolasdizzNo ratings yet
- Neural Correlates of Consciousness - Koch 2016Document15 pagesNeural Correlates of Consciousness - Koch 2016Juan Cristobal Ruiz Diaz100% (1)
- Autopoiesis PDFDocument44 pagesAutopoiesis PDFnicolasdizzNo ratings yet
- Consciousness: Here, There and Everywhere?: ReviewDocument18 pagesConsciousness: Here, There and Everywhere?: ReviewnicolasdizzNo ratings yet
- NIH Public Access: Individual Variability in Functional Connectivity Architecture of The Human BrainDocument20 pagesNIH Public Access: Individual Variability in Functional Connectivity Architecture of The Human BrainnicolasdizzNo ratings yet
- 1b. Husserl - Cartesian Meditations (Introduction, Meditation 1)Document6 pages1b. Husserl - Cartesian Meditations (Introduction, Meditation 1)nicolasdizzNo ratings yet
- 1b. Husserl - Cartesian Meditations (Introduction, Meditation 1)Document6 pages1b. Husserl - Cartesian Meditations (Introduction, Meditation 1)nicolasdizzNo ratings yet
- 10 1523@jneurosci 3659-14 2015 PDFDocument8 pages10 1523@jneurosci 3659-14 2015 PDFnicolasdizzNo ratings yet
- Foetal and Neonatal Intracranial Haemorrhage in Term Newborn Infants: Hacettepe University ExperienceDocument6 pagesFoetal and Neonatal Intracranial Haemorrhage in Term Newborn Infants: Hacettepe University ExperiencenicolasdizzNo ratings yet
- Management and Predictors in Patients With Spontaneous Intracerebral HemorrhageDocument10 pagesManagement and Predictors in Patients With Spontaneous Intracerebral HemorrhagenicolasdizzNo ratings yet
- Emotion PDFDocument13 pagesEmotion PDFnicolasdizzNo ratings yet
- Management and Predictors in Patients With Spontaneous Intracerebral HemorrhageDocument10 pagesManagement and Predictors in Patients With Spontaneous Intracerebral HemorrhagenicolasdizzNo ratings yet
- LeaP Science 10 Q4 W3 4Document5 pagesLeaP Science 10 Q4 W3 4D Garcia100% (2)
- UltraSonic Processors - CatalogDocument2 pagesUltraSonic Processors - Catalogdéborah_rosalesNo ratings yet
- Class XI Half SlybussDocument10 pagesClass XI Half SlybussDevansh AgarwalNo ratings yet
- The Truth About Honey - White Paper PDFDocument5 pagesThe Truth About Honey - White Paper PDFSergio PérezNo ratings yet
- Science Paper 7 - SolvedDocument10 pagesScience Paper 7 - SolvedRavi KumarNo ratings yet
- Leveraging Big Data To Transform TargetDocument13 pagesLeveraging Big Data To Transform Targetdeweyev699No ratings yet
- An Examination and Critique of Current Methods To Determine Exercise IntensityDocument28 pagesAn Examination and Critique of Current Methods To Determine Exercise IntensityPabloAñonNo ratings yet
- Taxonomy of Haematococcus PluvialisDocument2 pagesTaxonomy of Haematococcus PluvialisKomathi BalasupramaniamNo ratings yet
- Accessory Organ of The AbdomenDocument58 pagesAccessory Organ of The AbdomenOgundipe olorunfemiNo ratings yet
- Kebutuhan Zat Gizi Untuk Pasien Pada Berbagai Gangguan: Oleh: Dr. Ikbal Gentar Alam, Mkes, SPGK, AifoDocument103 pagesKebutuhan Zat Gizi Untuk Pasien Pada Berbagai Gangguan: Oleh: Dr. Ikbal Gentar Alam, Mkes, SPGK, AifoRafsya JulwandikaNo ratings yet
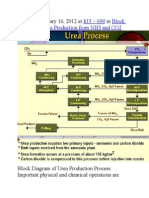
- Published January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2Document9 pagesPublished January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2himanshuchawla654No ratings yet
- Cat ExamDocument25 pagesCat Examlahsivlahsiv684No ratings yet
- Kaid Seloua-PdbDocument9 pagesKaid Seloua-Pdbseloua kaidNo ratings yet
- Ovulation CalanderDocument2 pagesOvulation CalanderBN NGNo ratings yet
- Recent DevelopmentsDocument295 pagesRecent DevelopmentsGabriel PinheiroNo ratings yet
- Manage Your Personal Energy PDFDocument46 pagesManage Your Personal Energy PDFezhil manickamNo ratings yet
- Chest TubesDocument34 pagesChest TubesMuhd ShafiqNo ratings yet
- 〈89.1〉 Collagenase IDocument5 pages〈89.1〉 Collagenase IAvijit HazraNo ratings yet
- Phet - Activity 1Document4 pagesPhet - Activity 1api-306913723No ratings yet
- Why Do Giraffes Have Long Necks?Document22 pagesWhy Do Giraffes Have Long Necks?johnosborneNo ratings yet
- Penilaian Sumatif Satuan Pendidikan (PSSP) Mata Pelajaran Bahasa Inggris Ta. 20222023Document1 pagePenilaian Sumatif Satuan Pendidikan (PSSP) Mata Pelajaran Bahasa Inggris Ta. 20222023Malfa FaradisNo ratings yet
- Final Project Report Kiran MazumdarDocument31 pagesFinal Project Report Kiran MazumdarPrajot Morajkar100% (3)
- Staar Eoc 2016test Bio F 7Document39 pagesStaar Eoc 2016test Bio F 7api-293216402No ratings yet
- Tule Elk in Point ReyesDocument4 pagesTule Elk in Point ReyesPointReyesNo ratings yet
- ALB2Document4 pagesALB2Jonalyn SalandoNo ratings yet
- Jane Goodall BiographyDocument2 pagesJane Goodall BiographyOjas JangalNo ratings yet
- All CompleteDocument10 pagesAll CompleteNanep NanenanenaNo ratings yet
- Short Note Biology Form 5-Chapter 3 Coordination and ResponseDocument6 pagesShort Note Biology Form 5-Chapter 3 Coordination and Responsesalamah_sabri75% (4)
- MoPSE Science Module Volume 1 - FINAL4WEBDocument336 pagesMoPSE Science Module Volume 1 - FINAL4WEBSibusiso Ntomby NkalaNo ratings yet