You might also like
- The DAMA Guide To The Data Management Body of Knowledge - First EditionDocument430 pagesThe DAMA Guide To The Data Management Body of Knowledge - First Editionkakarotodesu100% (10)
- Final Exam Tle Grade 8Document4 pagesFinal Exam Tle Grade 8John leo Claus67% (3)
- Hermes - GodDocument19 pagesHermes - GodAvocadoraNo ratings yet
- Eating Well: Snacks For 1-4 Year OldsDocument42 pagesEating Well: Snacks For 1-4 Year OldsAndreea AndreiNo ratings yet
- ScienceDocument4 pagesScienceBhargavi VeppalaNo ratings yet
- 1991 BP Nazareth EtalDocument6 pages1991 BP Nazareth EtalholmessbNo ratings yet
- Structural Studies of A Calmodulin Mutant With Defective Regulation of Muscle Contraction John Emmons and Adina KilpatrickDocument10 pagesStructural Studies of A Calmodulin Mutant With Defective Regulation of Muscle Contraction John Emmons and Adina KilpatrickzaxNo ratings yet
- A Calcium Release Activated Calcium Influx in Primary Cultures of Rat Osteoblast-Like CellsDocument6 pagesA Calcium Release Activated Calcium Influx in Primary Cultures of Rat Osteoblast-Like CellsJeremy BellNo ratings yet
- Manoalide, Natural Sesterterpenoid That Inhibits Calcium Channels"Document8 pagesManoalide, Natural Sesterterpenoid That Inhibits Calcium Channels"Collin LoganNo ratings yet
- Aguilar 2002 LCRDocument5 pagesAguilar 2002 LCRiplabaNo ratings yet
- Nucleic: Efficiency E.coli by ElectroporationDocument19 pagesNucleic: Efficiency E.coli by Electroporationh_spriyaNo ratings yet
- Coulton1982 FhuADocument9 pagesCoulton1982 FhuAhugoNo ratings yet
- Characterization of A Epilactose-Productiong Cellobiose 2-Epimerase From Thermoanaerobacterium SacharolyticumDocument6 pagesCharacterization of A Epilactose-Productiong Cellobiose 2-Epimerase From Thermoanaerobacterium SacharolyticumKukymovNo ratings yet
- Flow Calcium Kinetics Chapter15Document5 pagesFlow Calcium Kinetics Chapter15Marica GemeiNo ratings yet
- 0 GlucagonDocument12 pages0 GlucagonRenata Lima de FreitasNo ratings yet
- V. Sanz-Nebot Et Al. J. Chromatogr. B 798 2003Document7 pagesV. Sanz-Nebot Et Al. J. Chromatogr. B 798 2003bookmoonNo ratings yet
- Electrochemical Bienzyme Membrane Sensor For Free CholesterolDocument11 pagesElectrochemical Bienzyme Membrane Sensor For Free CholesterolAnand ChelliahNo ratings yet
- 89 JBC - 264 - 17309Document7 pages89 JBC - 264 - 17309Gregory BurrowsNo ratings yet
- PDFDocument11 pagesPDFAstridz Monsteqszia SilvanaNo ratings yet
- 09 Chapter 4Document33 pages09 Chapter 4dgdNo ratings yet
- Purif ACE G5Document9 pagesPurif ACE G5simon.ignacio.jNo ratings yet
- 2 CloningDocument7 pages2 CloningMiguel MirandaNo ratings yet
- Ja8b11449 Si 001Document29 pagesJa8b11449 Si 001Angel Ernesto Perales RochaNo ratings yet
- TMP 633 EDocument5 pagesTMP 633 EFrontiersNo ratings yet
- Calcium Dependence of Calcium Extrusion and Calcium Uptake in Mouse Pancreatic Acinar Cells J Physiol 1996Document9 pagesCalcium Dependence of Calcium Extrusion and Calcium Uptake in Mouse Pancreatic Acinar Cells J Physiol 1996Pedro J CamelloNo ratings yet
- Escherichia Colz K12Document5 pagesEscherichia Colz K12RenitaNo ratings yet
- 1423 FullDocument12 pages1423 Fullashish_jnuNo ratings yet
- Paper 7Document10 pagesPaper 7axe123anishNo ratings yet
- Rolul Taurinei in SNCDocument6 pagesRolul Taurinei in SNCMihaela ȚarnăNo ratings yet
- Simultaneous Monitoring of Glucose, Lactate, - Glutamate and Hypoxanthine Levels in Rat Striatum by A Ow-Injection Enzyme Electrode Array System With in Vivo Microdialysis SamplingDocument7 pagesSimultaneous Monitoring of Glucose, Lactate, - Glutamate and Hypoxanthine Levels in Rat Striatum by A Ow-Injection Enzyme Electrode Array System With in Vivo Microdialysis SamplingsggdgdNo ratings yet
- Ca2+ Homeostasis in The Endoplasmic Reticulum Measured With A NewDocument9 pagesCa2+ Homeostasis in The Endoplasmic Reticulum Measured With A Newser5885No ratings yet
- Interaction of Some Local Anaesthetics and Antisickling Drugs With Calcium-Carrier Proteins in BiomenbranesDocument8 pagesInteraction of Some Local Anaesthetics and Antisickling Drugs With Calcium-Carrier Proteins in BiomenbranesClement BewajiNo ratings yet
- Meissner 2008Document13 pagesMeissner 2008Farhana AnuarNo ratings yet
- Escherichia Coli: Two Genetically-Distinct and Differentially Regulated Aconitases (Acna and Acnb)Document11 pagesEscherichia Coli: Two Genetically-Distinct and Differentially Regulated Aconitases (Acna and Acnb)sgouni123No ratings yet
- Elektroforesis KapilerDocument4 pagesElektroforesis KapilerRachma DyraNo ratings yet
- Mitochondrial Ca2+ Concentrations in Live CellsDocument14 pagesMitochondrial Ca2+ Concentrations in Live Cellsser5885No ratings yet
- Zeta Potential Mediated Reaction Monitoring On Nano and MicroparticlesDocument7 pagesZeta Potential Mediated Reaction Monitoring On Nano and Microparticlest3guhku2hNo ratings yet
- Menadione PKDocument7 pagesMenadione PKmadremNo ratings yet
- Expression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedDocument0 pagesExpression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedMaísa FonsecaNo ratings yet
- Electrical Characters On Surface of Apoptotic Cell in Maize Roots Induced by CytotoxinsDocument10 pagesElectrical Characters On Surface of Apoptotic Cell in Maize Roots Induced by CytotoxinsUmiatin RamdhaniNo ratings yet
- Immunological Cross-Reactivity Between Calcium ATPases From Human and Porcine ErythrocytesDocument7 pagesImmunological Cross-Reactivity Between Calcium ATPases From Human and Porcine ErythrocytesClement BewajiNo ratings yet
- Cyclic AMP-mediated Inhibition of Noradrenaline-Induced Contraction Ca Influx in Guinea-Pig Vas DeferensDocument12 pagesCyclic AMP-mediated Inhibition of Noradrenaline-Induced Contraction Ca Influx in Guinea-Pig Vas DeferensssmorgNo ratings yet
- Microchemical Journal: 3 4 2 4 Wiem Bouali, Nevin Erk, Asena Ayse Genc, Hassan Elzain Hassan AHMED, Mustafa SoylakDocument9 pagesMicrochemical Journal: 3 4 2 4 Wiem Bouali, Nevin Erk, Asena Ayse Genc, Hassan Elzain Hassan AHMED, Mustafa Soylakhassanelzain84No ratings yet
- LT2 - Modelo Cinetico Pseudo-Intrinseco de La Bioconversion de CetonaDocument9 pagesLT2 - Modelo Cinetico Pseudo-Intrinseco de La Bioconversion de CetonaMafer CVNo ratings yet
- Polymerase Chain Reaction: Determining The Right Magnesium Chloride Concentration For DNA Amplification of Acetobacter XylinumDocument5 pagesPolymerase Chain Reaction: Determining The Right Magnesium Chloride Concentration For DNA Amplification of Acetobacter XylinumAlyza Joy RamirezNo ratings yet
- Journal of Bacteriology-1984-Matsumura-413.fullDocument8 pagesJournal of Bacteriology-1984-Matsumura-413.fullthumita kumiNo ratings yet
- Advances in The PurificationDocument10 pagesAdvances in The Purificationsantanaerik2005No ratings yet
- Uptake of Transfer Ribonucleic and Leukemic Cells : Acid NormalDocument8 pagesUptake of Transfer Ribonucleic and Leukemic Cells : Acid NormalianjprNo ratings yet
- Cabana 2007Document9 pagesCabana 2007Valeria MirandaNo ratings yet
- A Prodrug Approach For Improving Antituberculosis Activity of PotentDocument22 pagesA Prodrug Approach For Improving Antituberculosis Activity of PotentPhablo vinicius dos santos carneiroNo ratings yet
- El Rol Del Sensor de CalcioDocument10 pagesEl Rol Del Sensor de CalcioLeandro QuitoNo ratings yet
- Research Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsDocument12 pagesResearch Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsCarlos LabradaNo ratings yet
- Stoichiometry: Calcium/sodiumDocument6 pagesStoichiometry: Calcium/sodiumIvan CirkovicNo ratings yet
- Art 1Document18 pagesArt 1Luis Hurtado PardoNo ratings yet
- Aerts (2008) CMLS 65, 1933-Ydc1p Ceramidase Triggers Organelle Fragmentation, Apoptosis and Aging in YeastsDocument10 pagesAerts (2008) CMLS 65, 1933-Ydc1p Ceramidase Triggers Organelle Fragmentation, Apoptosis and Aging in YeastsDario FernándezNo ratings yet
- HCV ActivityDocument8 pagesHCV ActivityDr. P. Ramu SridharNo ratings yet
- Electrical Stress-Directed Evolution of Biocatalyst Texcoco Soil Community For Microbial Fuel CellDocument9 pagesElectrical Stress-Directed Evolution of Biocatalyst Texcoco Soil Community For Microbial Fuel CellKamaraj Sathish KumarNo ratings yet
- Isolation and Purification of CellulaseDocument6 pagesIsolation and Purification of CellulaseMeshal NoorNo ratings yet
- Full Paper Novel Sulphamides and Sulphonamides Incorporating The Tetralin Scaffold As Carbonic Anhydrase and Acetylcholine Esterase InhibitorsDocument9 pagesFull Paper Novel Sulphamides and Sulphonamides Incorporating The Tetralin Scaffold As Carbonic Anhydrase and Acetylcholine Esterase InhibitorsRajesh KumarNo ratings yet
- Gentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsDocument0 pagesGentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsCLPHtheoryNo ratings yet
- Membrane Proteins: FEBS Federation of European Biochemical Societies: 11th Meeting, Copenhagen, 1977From EverandMembrane Proteins: FEBS Federation of European Biochemical Societies: 11th Meeting, Copenhagen, 1977No ratings yet
- Transition Metal Catalyzed Furans Synthesis: Transition Metal Catalyzed Heterocycle Synthesis SeriesFrom EverandTransition Metal Catalyzed Furans Synthesis: Transition Metal Catalyzed Heterocycle Synthesis SeriesNo ratings yet
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967From EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterNo ratings yet
- Toxicidad TimerosalDocument45 pagesToxicidad TimerosalIta Zuñiga AlfaroNo ratings yet
- Caminar Reduce Los AVE en HombreDocument9 pagesCaminar Reduce Los AVE en HombreIta Zuñiga AlfaroNo ratings yet
- 31 Humphrey Mitochondrial Initiated Thimerosal 2005Document10 pages31 Humphrey Mitochondrial Initiated Thimerosal 2005Ita Zuñiga AlfaroNo ratings yet
- Measles Rubella Monitoring 170424Document16 pagesMeasles Rubella Monitoring 170424Ita Zuñiga AlfaroNo ratings yet
- 9 Donoso 2000Document8 pages9 Donoso 2000Ita Zuñiga AlfaroNo ratings yet
- 11 Montero 2001Document10 pages11 Montero 2001Ita Zuñiga AlfaroNo ratings yet
- 9 Donoso 2000Document8 pages9 Donoso 2000Ita Zuñiga AlfaroNo ratings yet
- 9 Donoso 2000Document8 pages9 Donoso 2000Ita Zuñiga AlfaroNo ratings yet
- 5 Anundi 1979 Glutathione Depletion in Isolated Hepatocytes1Document7 pages5 Anundi 1979 Glutathione Depletion in Isolated Hepatocytes1Ita Zuñiga AlfaroNo ratings yet
- Docs Grails Org 4 0 0 Guide Single HTMLDocument296 pagesDocs Grails Org 4 0 0 Guide Single HTMLlicface13No ratings yet
- With Pneumatic and Electric Actuators: Datasheet 448001 EnglishDocument7 pagesWith Pneumatic and Electric Actuators: Datasheet 448001 EnglishPinak ProjectsNo ratings yet
- Presentation 1Document11 pagesPresentation 1CJ CastroNo ratings yet
- A 268 - A 268M - 01 Qti2oc0wmq - PDFDocument6 pagesA 268 - A 268M - 01 Qti2oc0wmq - PDFMan98No ratings yet
- Department of Labor: BC Bond ListDocument67 pagesDepartment of Labor: BC Bond ListUSA_DepartmentOfLabor100% (1)
- Inertia Physics: Defi Ni Ti OnDocument2 pagesInertia Physics: Defi Ni Ti OnSentoash NaiduNo ratings yet
- Output D4Document34 pagesOutput D4Angel MingaNo ratings yet
- Segment Reporting Decentralized Operations and Responsibility Accounting SystemDocument34 pagesSegment Reporting Decentralized Operations and Responsibility Accounting SystemalliahnahNo ratings yet
- Anti LeproticDocument9 pagesAnti LeproticMeenakshi shARMANo ratings yet
- Chapter 12 Fraud and ErrorsDocument3 pagesChapter 12 Fraud and ErrorsMajoy BantocNo ratings yet
- Initial 2Document6 pagesInitial 2Asad HoseinyNo ratings yet
- Physics 715 HW 1Document13 pagesPhysics 715 HW 1Antonildo PereiraNo ratings yet
- Airport Lounges Industry-Report-Frost-SullivanDocument112 pagesAirport Lounges Industry-Report-Frost-SullivansandeepNo ratings yet
- RLA-Grade 6 2023Document4 pagesRLA-Grade 6 2023Catherine Mabini BeatoNo ratings yet
- New Criticism Hills Like White Elephants FinalDocument4 pagesNew Criticism Hills Like White Elephants Finalapi-313631761No ratings yet
- EKOFLUID PRODUCT SHEET FILOIL 6000 210x297 EN WEBDocument4 pagesEKOFLUID PRODUCT SHEET FILOIL 6000 210x297 EN WEBjunaidi.036No ratings yet
- Air Quality StandardsDocument2 pagesAir Quality StandardsJanmejaya BarikNo ratings yet
- Securing Your Organization From Modern Ransomware: Ransomware Attacks Are Now A Team EffortDocument11 pagesSecuring Your Organization From Modern Ransomware: Ransomware Attacks Are Now A Team EfforttiagouebemoraisNo ratings yet
- Activate My Win 7 Pro Product KeyDocument4 pagesActivate My Win 7 Pro Product KeydoktauNo ratings yet
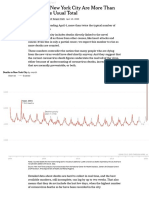
- Deaths in New York City Are More Than Double The Usual TotalDocument3 pagesDeaths in New York City Are More Than Double The Usual TotalRamón RuizNo ratings yet
- Capital MarketDocument5 pagesCapital MarketBalamanichalaBmcNo ratings yet
- Aurora National High School: Report On AttendanceDocument2 pagesAurora National High School: Report On AttendanceLimuel CaringalNo ratings yet
- Mat210 LectureNotes 1Document7 pagesMat210 LectureNotes 1Franch Maverick Arellano LorillaNo ratings yet
- S.NO. Students Name Contact Number CAP 10TH: Ideal Institute of Pharamacy, Wada SESSION-2020-21Document12 pagesS.NO. Students Name Contact Number CAP 10TH: Ideal Institute of Pharamacy, Wada SESSION-2020-21DAMBALENo ratings yet
- Polyethylene PolyamineDocument6 pagesPolyethylene PolyamineAV kayanNo ratings yet
- Tahmina Ferdousy Jhumu: HND Btec Unit 15 Psychology For Health and Social CareDocument29 pagesTahmina Ferdousy Jhumu: HND Btec Unit 15 Psychology For Health and Social CareNabi BoxNo ratings yet