You might also like
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5813)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Crop ScienceDocument66 pagesCrop ScienceLaila UbandoNo ratings yet
- Nitric Acid - MTI MS-5Document170 pagesNitric Acid - MTI MS-5rodrigomiyawakiNo ratings yet
- Baseline Corrosion SurveyDocument16 pagesBaseline Corrosion SurveyNguyen Anh Tung100% (7)
- Hold Time Study of 70% IpaDocument5 pagesHold Time Study of 70% IpaAshok Lenka100% (4)
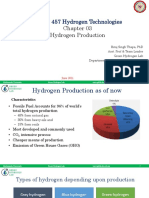
- MEPP 457 Hydrogen TechnologiesDocument88 pagesMEPP 457 Hydrogen TechnologiesAshim LamichhaneNo ratings yet
- Orbu SPC 44 0 22Document98 pagesOrbu SPC 44 0 22geverett2765No ratings yet
- Adsorption Process For Natural GAS TreatmentDocument24 pagesAdsorption Process For Natural GAS TreatmentAhmed ElShora100% (3)
- Optical MicrosDocument73 pagesOptical MicrosSaurabhBhardwajNo ratings yet
- Typical Material Specifications For Major Component PartsDocument9 pagesTypical Material Specifications For Major Component PartsSampath KumarNo ratings yet
- The Synthesis of Water Soluble N-Acyl Chitosan Derivatives For CHDocument126 pagesThe Synthesis of Water Soluble N-Acyl Chitosan Derivatives For CHAmtoni Cesar NainggolanNo ratings yet
- CLG Isocracking Tech SheetDocument2 pagesCLG Isocracking Tech Sheetkrishna100% (1)
- 01-12-19 Sri Chaitanya-Sr - Chaina-I - L-I&II - Jee-Adv (2017-P2) - PTA-12 - Q.PDocument19 pages01-12-19 Sri Chaitanya-Sr - Chaina-I - L-I&II - Jee-Adv (2017-P2) - PTA-12 - Q.PSridhar ReddyNo ratings yet
- Polyatomic IonsDocument2 pagesPolyatomic IonsJuan KorNo ratings yet
- Textile DyesDocument5 pagesTextile Dyesapi-27596299No ratings yet
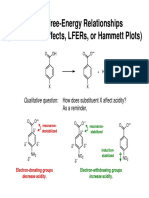
- Linear Free Energy RelationshipDocument16 pagesLinear Free Energy RelationshipAb IrizarryNo ratings yet
- CHAPTER III Blood and Blood StainsDocument11 pagesCHAPTER III Blood and Blood StainsArchie ToribioNo ratings yet
- Water-in-Crude Oil EmulsionDocument30 pagesWater-in-Crude Oil EmulsionMsd KrishnaNo ratings yet
- Biomaterials Lecture #3 Notes AmalgamDocument3 pagesBiomaterials Lecture #3 Notes AmalgamZ FNo ratings yet
- Lightcraft PropulsionDocument37 pagesLightcraft PropulsionLin CornelisonNo ratings yet
- Cell Respiration Cornell Notes TeacherDocument5 pagesCell Respiration Cornell Notes TeacherErik Artur100% (1)
- Cocoa Beans Shelf LifeDocument1 pageCocoa Beans Shelf Lifeibrail5No ratings yet
- 01 Water Suppression SystemDocument54 pages01 Water Suppression SystemKolos Char YiuNo ratings yet
- Effects of Gamma On Medical DevicesDocument7 pagesEffects of Gamma On Medical DevicesLin ZinNo ratings yet
- 05a Qualitative Analysis For Organic CompoundsDocument5 pages05a Qualitative Analysis For Organic CompoundsReyo VillanuevaNo ratings yet
- Allen: Aieee - 2012 Test Paper With Answer (Held On Sunday 29 APRIL, 2012)Document5 pagesAllen: Aieee - 2012 Test Paper With Answer (Held On Sunday 29 APRIL, 2012)rishabhk28995No ratings yet
- 1.1 Traced Lines: Fig 1.1 - Insulating Tracer and Product LinesDocument14 pages1.1 Traced Lines: Fig 1.1 - Insulating Tracer and Product Linesmurugn08No ratings yet
- Central Dogma of BiologyDocument28 pagesCentral Dogma of BiologyJabin Sta. TeresaNo ratings yet
- Yearwise Chem ESA PapersDocument29 pagesYearwise Chem ESA PapersGagan SL100% (1)
- Mobil DTE - 2Document10 pagesMobil DTE - 2sasikumartvsNo ratings yet