You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 10k Training Program (Interval, Cruise and Long Dist)Document6 pages10k Training Program (Interval, Cruise and Long Dist)kevinalmerNo ratings yet
- Obstructive Disease of Lung and Physiotherapy ManagementDocument76 pagesObstructive Disease of Lung and Physiotherapy Managementphysio43100% (1)
- FRACP Cardiology Study Notes Volume 1Document543 pagesFRACP Cardiology Study Notes Volume 1not here 2make friends sorryNo ratings yet
- Heterogeneous Hunter-Gatherer and SteppeRelated Ancestries in Late Neolithic and Bell Beaker Genomes From Present-Day FranceDocument23 pagesHeterogeneous Hunter-Gatherer and SteppeRelated Ancestries in Late Neolithic and Bell Beaker Genomes From Present-Day FranceJan KowalskiNo ratings yet
- Oι σημερινοί Έλληνες όμοιοι με τους πληθυσμούς του Β. Αιγαίου του 2.000 π.Χ.Document44 pagesOι σημερινοί Έλληνες όμοιοι με τους πληθυσμούς του Β. Αιγαίου του 2.000 π.Χ.ARISTEIDIS VIKETOSNo ratings yet
- Lazaridis, I. Genetic Origins of The Minoans and MycenaeansDocument17 pagesLazaridis, I. Genetic Origins of The Minoans and MycenaeansRenan Falcheti Peixoto100% (3)
- Neolithic Genomic Data From Southern France Showcase Intensified Interactions With Huntergatherer CommunitiesDocument28 pagesNeolithic Genomic Data From Southern France Showcase Intensified Interactions With Huntergatherer CommunitiesJan KowalskiNo ratings yet
- Ancient Plagues Uncovered in Bronze Age CreteDocument23 pagesAncient Plagues Uncovered in Bronze Age CreteJan KowalskiNo ratings yet
- The Diverse Genetic Origins of A Classical Period Greek ArmyDocument11 pagesThe Diverse Genetic Origins of A Classical Period Greek ArmyJan KowalskiNo ratings yet
- Population History of Early Medieval UkraineDocument1 pagePopulation History of Early Medieval UkraineJan KowalskiNo ratings yet
- Early Bronze Age Aegean Genomes From Perachora Cave, Corinth, GreeceDocument1 pageEarly Bronze Age Aegean Genomes From Perachora Cave, Corinth, GreeceJan KowalskiNo ratings yet
- Neolithization of North Africa Involved The Migration of 2 People From Both The Levant and EuropeDocument14 pagesNeolithization of North Africa Involved The Migration of 2 People From Both The Levant and EuropeJan KowalskiNo ratings yet
- An Admixture Signal in Armenians Around The End of The Bronze Age Reveals Widespread Population Movement Across The Middle EastDocument19 pagesAn Admixture Signal in Armenians Around The End of The Bronze Age Reveals Widespread Population Movement Across The Middle EastJan KowalskiNo ratings yet
- Ancient DNA Reveals Admixture History and Endogamy in The Prehistoric AegeanDocument25 pagesAncient DNA Reveals Admixture History and Endogamy in The Prehistoric AegeanJan KowalskiNo ratings yet
- Late Danubian Mitochondrial Genomes Shed Light Into The Neolithisation of Central Europe in The 5th Millennium BCDocument12 pagesLate Danubian Mitochondrial Genomes Shed Light Into The Neolithisation of Central Europe in The 5th Millennium BCJan KowalskiNo ratings yet
- Genetic Insights Into The Social Organisation of The Avar Period Elite in The 7th Century AD Carpathian BasinDocument14 pagesGenetic Insights Into The Social Organisation of The Avar Period Elite in The 7th Century AD Carpathian BasinJan KowalskiNo ratings yet
- Genome-Wide Data From Two Early Neolithic East Asian Individuals Dating To 7700 Years AgoDocument11 pagesGenome-Wide Data From Two Early Neolithic East Asian Individuals Dating To 7700 Years AgoJan KowalskiNo ratings yet
- Pleistocene North African Genomes Link Near Eastern and Sub-Saharan African Human PopulationsDocument13 pagesPleistocene North African Genomes Link Near Eastern and Sub-Saharan African Human PopulationsJan KowalskiNo ratings yet
- The Genetic Makings of South AsiaDocument6 pagesThe Genetic Makings of South AsiaJan KowalskiNo ratings yet
- Ancient DNA Reveals Matrilineal Continuity in PresentDay Poland Over The Last Two MillenniaDocument9 pagesAncient DNA Reveals Matrilineal Continuity in PresentDay Poland Over The Last Two MillenniaJan Kowalski100% (1)
- Molecular Phylogeography of Armenians Based On Complete Mitochondrial Genome DataDocument113 pagesMolecular Phylogeography of Armenians Based On Complete Mitochondrial Genome DataJan KowalskiNo ratings yet
- Between The Baltic and Danubian Worlds: The Genetic Affinities of A Middle Neolithic Population From Central PolandDocument17 pagesBetween The Baltic and Danubian Worlds: The Genetic Affinities of A Middle Neolithic Population From Central PolandJan KowalskiNo ratings yet
- Genomic Analyses of Pre-European Conquest Human Remains From The Canary Islands Reveal Close Affinity To Modern North AfricansDocument13 pagesGenomic Analyses of Pre-European Conquest Human Remains From The Canary Islands Reveal Close Affinity To Modern North AfricansJan KowalskiNo ratings yet
- Ancient DNA Suggests Steppe Migrations Spread Indo-European LanguagesDocument17 pagesAncient DNA Suggests Steppe Migrations Spread Indo-European LanguagesJan KowalskiNo ratings yet
- Poster Scythians MariJarveDocument1 pagePoster Scythians MariJarveJan KowalskiNo ratings yet
- Extensive Farming in Estonia Started Through A Sex-Biased Migration From The SteppeDocument36 pagesExtensive Farming in Estonia Started Through A Sex-Biased Migration From The SteppeJan KowalskiNo ratings yet
- Investigating Kinship of Neolithic post-LBK Human Remains From Krusza Zamkowa, Poland Using Ancient DNADocument10 pagesInvestigating Kinship of Neolithic post-LBK Human Remains From Krusza Zamkowa, Poland Using Ancient DNAJan KowalskiNo ratings yet
- Investigating Kinship of Neolithic post-LBK Human Remains From Krusza Zamkowa, Poland Using Ancient DNADocument10 pagesInvestigating Kinship of Neolithic post-LBK Human Remains From Krusza Zamkowa, Poland Using Ancient DNAJan KowalskiNo ratings yet
- S1 Appendix 2 The Archaeological Background of Current Study in EstoniaDocument17 pagesS1 Appendix 2 The Archaeological Background of Current Study in EstoniaJan KowalskiNo ratings yet
- Discussion Are The Origins of Indo-EuropDocument15 pagesDiscussion Are The Origins of Indo-EuropChetanNo ratings yet
- Genome-Wide Data From Two Early Neolithic East Asian Individuals Dating To 7700 Years AgoDocument11 pagesGenome-Wide Data From Two Early Neolithic East Asian Individuals Dating To 7700 Years AgoJan KowalskiNo ratings yet
- Genomics of Mesolithic Scandinavia Reveal Colonization Routes and Highlatitude AdaptationDocument14 pagesGenomics of Mesolithic Scandinavia Reveal Colonization Routes and Highlatitude AdaptationJan KowalskiNo ratings yet
- The Physiologic Nature of SleepDocument660 pagesThe Physiologic Nature of SleepRaluca HuzumNo ratings yet
- Topic 3 NCM 112Document4 pagesTopic 3 NCM 112Marielle ChuaNo ratings yet
- Aakash Zoology Study Package 3 SolutionsssssssDocument104 pagesAakash Zoology Study Package 3 SolutionsssssssRishika PaulNo ratings yet
- Digestion and absorption multiple choice questionsDocument46 pagesDigestion and absorption multiple choice questionsAmbika dawadiNo ratings yet
- The Effects of Shade Levels and Additional Calcium Nitrate On The Morphology of Cyrtandra Bracheia B.L. Burt (Siti Maslizah) PP 18-25Document8 pagesThe Effects of Shade Levels and Additional Calcium Nitrate On The Morphology of Cyrtandra Bracheia B.L. Burt (Siti Maslizah) PP 18-25upenapahangNo ratings yet
- British J Pharmacology - 2021 - ZobdehDocument20 pagesBritish J Pharmacology - 2021 - ZobdehRenju KuriakoseNo ratings yet
- Cervical SpineDocument69 pagesCervical Spineveravero100% (3)
- Tamponade KordisDocument20 pagesTamponade KordisMuhammad Aprianto RamadhanNo ratings yet
- CHE 475 Biochemical Engineering II: Course SyllabusDocument19 pagesCHE 475 Biochemical Engineering II: Course SyllabusAkib Atique Khan DhruboNo ratings yet
- Rhinologic and Sleep Apnea Surgical Techniques 1st Ed 2007 SDocument427 pagesRhinologic and Sleep Apnea Surgical Techniques 1st Ed 2007 SKarolina RamirezNo ratings yet
- Annas Test ResultsDocument1 pageAnnas Test Resultsapi-307430259No ratings yet
- Ch.11. Transportation in Animals and PlantsDocument3 pagesCh.11. Transportation in Animals and PlantsSharath ChandraNo ratings yet
- Results of Analysis - Kae Kru ArtDocument3 pagesResults of Analysis - Kae Kru ArtDr. Siraphop OneSongchai RatanasubanNo ratings yet
- (FRANCISCO) Activity 4.2Document1 page(FRANCISCO) Activity 4.2Kyle Isabelle FranciscoNo ratings yet
- PDF 1.1 Shoulder-MinDocument5 pagesPDF 1.1 Shoulder-MinGhea Putri HendrianiNo ratings yet
- Gas Exchange in the Lungs - How Oxygen and Carbon Dioxide are TransferredDocument15 pagesGas Exchange in the Lungs - How Oxygen and Carbon Dioxide are TransferredThe Deep Sea IdNo ratings yet
- Gayatri Gaikwad (Biology)Document29 pagesGayatri Gaikwad (Biology)prashik vigheNo ratings yet
- Range of MotionDocument17 pagesRange of MotionJim Cariaga100% (2)
- New - Biotechnology (Class XII)Document167 pagesNew - Biotechnology (Class XII)Rahul JaiswalNo ratings yet
- Vertebra Anatomy 125Document110 pagesVertebra Anatomy 125Jennifer FirestoneNo ratings yet
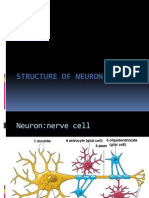
- Structure of NeuronDocument12 pagesStructure of NeuronMurali SambhuNo ratings yet
- Masaj Facial - EngDocument4 pagesMasaj Facial - EngLucian CosneanuNo ratings yet
- Vertical Root FractureDocument2 pagesVertical Root FractureJeremiah SchmidtNo ratings yet
- Pulmonary Function Test: By: Alfaro, Ruby Jane SDocument13 pagesPulmonary Function Test: By: Alfaro, Ruby Jane SgjevamNo ratings yet
- CNS Stimulants: Classification, Mechanisms and UsesDocument20 pagesCNS Stimulants: Classification, Mechanisms and UsesAn LoNo ratings yet
- Labreport V SMSH 23 10141161 PDFDocument2 pagesLabreport V SMSH 23 10141161 PDFMohanNo ratings yet
- KKYDocument44 pagesKKYAdm RajaNo ratings yet