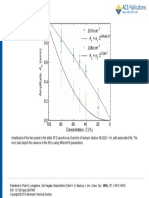
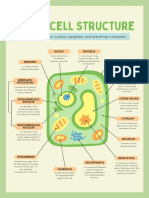
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Qgui ManualDocument39 pagesQgui ManualyNo ratings yet
- Butler 2018Document9 pagesButler 2018yNo ratings yet
- Book 2Document1 pageBook 2yNo ratings yet
- What You Need To Know About Python (Ebook) - Pierluigi RitiDocument66 pagesWhat You Need To Know About Python (Ebook) - Pierluigi RitiSelf-Developer75% (4)
- 10 1126@science Aat4010Document7 pages10 1126@science Aat4010yNo ratings yet
- Glycosidase MechanismsDocument8 pagesGlycosidase MechanismsyNo ratings yet
- Amber16 PDFDocument923 pagesAmber16 PDFyNo ratings yet
- 10 1021-Acs Jpclett 5b00281figure4Document1 page10 1021-Acs Jpclett 5b00281figure4yNo ratings yet
- 10 1021-Jacs 5b07845figure5Document1 page10 1021-Jacs 5b07845figure5yNo ratings yet
- READMEDocument1 pageREADMEyNo ratings yet
- 10 1021-Jacs 5b07845figure3Document1 page10 1021-Jacs 5b07845figure3yNo ratings yet
- 10 1021-Acs Jpclett 5b00281figure3Document1 page10 1021-Acs Jpclett 5b00281figure3yNo ratings yet
- Ja5b03945 Si 001Document37 pagesJa5b03945 Si 001yNo ratings yet
- 10 1021-Acs Jpclett 5b00281figure2Document1 page10 1021-Acs Jpclett 5b00281figure2yNo ratings yet
- 2pt Gromacs DocumentationDocument12 pages2pt Gromacs DocumentationyNo ratings yet
- File 1195Document86 pagesFile 1195yNo ratings yet
- 10 1021-Jacs 5b07845figure5Document1 page10 1021-Jacs 5b07845figure5yNo ratings yet
- LieDocument7 pagesLieyNo ratings yet
- Ultrasound Velocity in WaterDocument3 pagesUltrasound Velocity in WaterAndrea EspinosaNo ratings yet
- Scoring of Nine Ligands in P450camDocument5 pagesScoring of Nine Ligands in P450camyNo ratings yet
- File 1270Document42 pagesFile 1270yNo ratings yet
- C+Mathematica Comp PhysDocument299 pagesC+Mathematica Comp PhysyNo ratings yet
- Free Energy Perturbation AnalysisDocument12 pagesFree Energy Perturbation AnalysisyNo ratings yet
- Photosynthesis and Cellular Respiration SummaryDocument5 pagesPhotosynthesis and Cellular Respiration Summarylady ann jimenezNo ratings yet
- Introductory Activities: Case StudiesDocument16 pagesIntroductory Activities: Case StudiesormattNo ratings yet
- Chemical Pathology 2 - Plasma ProteinsDocument5 pagesChemical Pathology 2 - Plasma ProteinsaNo ratings yet
- Atherosclerosis: Process, Indicators, Risk Factors and New HopesDocument30 pagesAtherosclerosis: Process, Indicators, Risk Factors and New HopesSalwiyadiNo ratings yet
- Fqah0111 2008Document5 pagesFqah0111 2008Wan Muhammad FaizuddinNo ratings yet
- Revision Notes For Class 12 CBSE Chemistry, Biomolecules - TopperlearningDocument14 pagesRevision Notes For Class 12 CBSE Chemistry, Biomolecules - TopperlearningRishabh Bhandari100% (2)
- Experiment 8 and 9 PDFDocument17 pagesExperiment 8 and 9 PDFKrizzi Dizon GarciaNo ratings yet
- WORKSHEET 3.1 Movement of Subtances Across The Plasma MembraneDocument3 pagesWORKSHEET 3.1 Movement of Subtances Across The Plasma MembraneFadhliana UzalliNo ratings yet
- Cambridge International AS & A Level: Biology 9700/12Document16 pagesCambridge International AS & A Level: Biology 9700/12shabanaNo ratings yet
- About Basic Local Alignment Search ToolDocument17 pagesAbout Basic Local Alignment Search Toolsaswati01No ratings yet
- Exploring Tissue Architecture Using Spatial Transcriptomics - BiozionDocument25 pagesExploring Tissue Architecture Using Spatial Transcriptomics - Biozionalice Ann sajithNo ratings yet
- Laporan Praktikum Uji NinhidrinDocument10 pagesLaporan Praktikum Uji NinhidrinIndrie DwiraandaNo ratings yet
- Labeled ImmunoassaysDocument6 pagesLabeled ImmunoassaysAngelica Mae LasamNo ratings yet
- Alkaloids PDFDocument81 pagesAlkaloids PDFAmit88% (8)
- Summative Test #2 Gen BioDocument2 pagesSummative Test #2 Gen BioKennedy Fieldad Vagay100% (1)
- Molecular Systematic Lecture 1Document57 pagesMolecular Systematic Lecture 1hudaNo ratings yet
- Bio Cell Structure and Function Chart and Review PDFDocument27 pagesBio Cell Structure and Function Chart and Review PDFMuddihil MadzlanNo ratings yet
- Genbio 1 NotesDocument1 pageGenbio 1 NoteselishaNo ratings yet
- FirstChoice RLM-RACE Kit Manual AnnotatedDocument33 pagesFirstChoice RLM-RACE Kit Manual AnnotatedlandangeNo ratings yet
- Cell Wall Synthesis Inhibitors (CompleteDocument44 pagesCell Wall Synthesis Inhibitors (CompletegrayNo ratings yet
- Sarcoma de Ewing S NEJMDocument11 pagesSarcoma de Ewing S NEJMbanzethyskyNo ratings yet
- Araling Panlipunan 8 Quarter 1 Report CardDocument13 pagesAraling Panlipunan 8 Quarter 1 Report CardJade MillanteNo ratings yet
- Plant Cell Structure: Nucleus, Chloroplasts & MoreDocument1 pagePlant Cell Structure: Nucleus, Chloroplasts & MoreMackenzie ElyseNo ratings yet
- Phenylalanine and Tyrosine Metabolism (18 Oct)Document14 pagesPhenylalanine and Tyrosine Metabolism (18 Oct)api-19824406100% (6)
- Biomedical Importance Glycolysis Can Function Under Anaerobic ConditionsDocument5 pagesBiomedical Importance Glycolysis Can Function Under Anaerobic ConditionsCatalina Denise Blaquera FloresNo ratings yet
- CELS 191 Objective Based SelfDocument6 pagesCELS 191 Objective Based SelfMimi BoisterNo ratings yet
- Cell Biology Activity GuideDocument4 pagesCell Biology Activity GuidePamaran, Kristel DenishNo ratings yet
- Gene Therapy: Current Perspective and What The Future HoldsDocument10 pagesGene Therapy: Current Perspective and What The Future HoldskepegawaianNo ratings yet
- Microplate Readers BrochureDocument12 pagesMicroplate Readers BrochureJuan Paulo MorenoNo ratings yet
- Mark Schemes Edexcel Synoptic EssaysDocument9 pagesMark Schemes Edexcel Synoptic Essaysapi-3740691100% (1)