You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Mayan Zodiac Symbols and NamesDocument7 pagesMayan Zodiac Symbols and Namescstocksg150% (2)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Unisystem PiratesDocument8 pagesUnisystem PiratesAnime300100% (1)
- Biodiversity For Grade 6Document49 pagesBiodiversity For Grade 6Jennifer Silva-PabillareNo ratings yet
- Principles of Selection and Mating SystemsDocument29 pagesPrinciples of Selection and Mating SystemsUnknown StudentNo ratings yet
- Primary Survey & Secondary Survey: Presentor: Shellazianne Anak Ringam Date: 20 November 2015Document27 pagesPrimary Survey & Secondary Survey: Presentor: Shellazianne Anak Ringam Date: 20 November 2015Keluang Man WinchesterNo ratings yet
- Basic Anatomy and Physiology of Brain and Spinal CordDocument59 pagesBasic Anatomy and Physiology of Brain and Spinal CorddrmalikarifNo ratings yet
- Lynn's Sleepover v1Document18 pagesLynn's Sleepover v1Anonymous kTxhgUNo ratings yet
- WALIKI Mens Knits 2010Document17 pagesWALIKI Mens Knits 2010Waliki Design TeamNo ratings yet
- AMCAT CocubesDocument35 pagesAMCAT CocubesMuthyala Akhil100% (6)
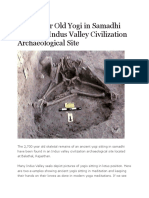
- 2,700 Year Old Yogi in Samadhi Found in Indus ValleyDocument12 pages2,700 Year Old Yogi in Samadhi Found in Indus ValleyMisterNo ratings yet
- Sample Proposal For GrantDocument8 pagesSample Proposal For GrantRomee SinghNo ratings yet
- Psychophysiological Detection of Deception Analysis Ii - Course #503Document67 pagesPsychophysiological Detection of Deception Analysis Ii - Course #503John PrasetioNo ratings yet
- Listening - Compare Animal and Human Behavior 2 - Mode - Report - Unit 6 - Lesson 3 - Vantage B2 - MyEnglishLabDocument1 pageListening - Compare Animal and Human Behavior 2 - Mode - Report - Unit 6 - Lesson 3 - Vantage B2 - MyEnglishLabAnnuar Florez100% (1)
- Vertical Root FractureDocument2 pagesVertical Root FractureJeremiah SchmidtNo ratings yet
- Endodontic Treatment FailureDocument8 pagesEndodontic Treatment FailureHawzheen SaeedNo ratings yet
- The Crane Frolics: The Path From Qihai To TanzhongDocument4 pagesThe Crane Frolics: The Path From Qihai To Tanzhongapi-268467409100% (1)
- TASK SHEET No. 1.2-1 Title: Perform Different Knots Used in Restraining Animals Performance ObjectiveDocument7 pagesTASK SHEET No. 1.2-1 Title: Perform Different Knots Used in Restraining Animals Performance ObjectiveRoh NaldzNo ratings yet
- DR Mushtaq Amna Ashraf Aqsa Bashir Ayesha Bibi Hira Nisar Iqra HabibDocument5 pagesDR Mushtaq Amna Ashraf Aqsa Bashir Ayesha Bibi Hira Nisar Iqra HabibtaibiNo ratings yet
- FOM STUDY GUIDE 3rd Block 1Document3 pagesFOM STUDY GUIDE 3rd Block 1Bernadine Cruz Par100% (1)
- Oxford Discover 2. SB. Transcripts Listening Sect. - 2019Document6 pagesOxford Discover 2. SB. Transcripts Listening Sect. - 2019Mon Abdel SalamNo ratings yet
- READ: Battle Creek Feral Cats OrdinanceDocument6 pagesREAD: Battle Creek Feral Cats OrdinanceJennifer BowmanNo ratings yet
- Chapter 7 Study GuideDocument4 pagesChapter 7 Study GuidejbradeeNo ratings yet
- D. L. Bliss State Park HandbookDocument4 pagesD. L. Bliss State Park HandbookCalifornia State ParksNo ratings yet
- ما يقارب 200 سؤال مع الاجابة لاسئلة الهيئة السعودية للتخصصات الصحية لاخصائيي المناعة والامصال وبDocument34 pagesما يقارب 200 سؤال مع الاجابة لاسئلة الهيئة السعودية للتخصصات الصحية لاخصائيي المناعة والامصال وبMohsen HaleemNo ratings yet
- Grade5 Unit1Document5 pagesGrade5 Unit1api-239494748No ratings yet
- African Grey Parrot - WikipediaDocument3 pagesAfrican Grey Parrot - WikipediaWilhelm Richard WagnerNo ratings yet
- Psychology of ThinkingDocument19 pagesPsychology of ThinkingDiyana HalimNo ratings yet
- Subcutaneous Infections and True PathogensDocument2 pagesSubcutaneous Infections and True PathogensthescarletpimpernelsNo ratings yet
- ĐỀ 48Document7 pagesĐỀ 48bichngocvonagiNo ratings yet
- I. EmbryologyDocument6 pagesI. EmbryologyEzra GenerosoNo ratings yet