You might also like
- Words Their Way Blue SortsDocument60 pagesWords Their Way Blue SortsSophie Lee75% (8)
- Step 1 UworldDocument25 pagesStep 1 UworldKarl Abiaad100% (23)
- Clinical Laboratory LawDocument26 pagesClinical Laboratory Lawlwlski71% (7)
- D-Values of PathogenDocument10 pagesD-Values of PathogenalirizalNo ratings yet
- International Journal of Food MicrobiologyDocument8 pagesInternational Journal of Food Microbiology0145093706No ratings yet
- The Acid Tolerance Response of Bacillus Cereus ATCC14579 Is Dependent On Culture PH, Growth Rate and Intracellular PHDocument11 pagesThe Acid Tolerance Response of Bacillus Cereus ATCC14579 Is Dependent On Culture PH, Growth Rate and Intracellular PHخديجة بيوNo ratings yet
- Exherichia Parte 2Document12 pagesExherichia Parte 2Gerson Moreno BermúdezNo ratings yet
- Breidt 2005 - Determination of 5-Log Pathogen Reduction TimesDocument6 pagesBreidt 2005 - Determination of 5-Log Pathogen Reduction TimespedroloxxxNo ratings yet
- Food MCB II NotesDocument73 pagesFood MCB II NotesRichard Simon KisituNo ratings yet
- JFP73 1304stanfordDocument9 pagesJFP73 1304stanfordDrSiva Prasad GuntamukkalaNo ratings yet
- Leyer & Johnson (1997)Document7 pagesLeyer & Johnson (1997)Kharisma N. PuspitasariNo ratings yet
- Bacillus Subtillis PHDocument10 pagesBacillus Subtillis PHanon_816002920No ratings yet
- Use of Acetic and Citric Acids To Inhibit Escherichia ColiDocument6 pagesUse of Acetic and Citric Acids To Inhibit Escherichia ColiRadwan AjoNo ratings yet
- Escherichia Coli O157:H7, Salmonella Enteritidis, and Listeria Monocytogenes On The Surfaces of TomatoesDocument9 pagesEscherichia Coli O157:H7, Salmonella Enteritidis, and Listeria Monocytogenes On The Surfaces of TomatoesRuL'zNo ratings yet
- Effects of Commercial Organic Acid Blends On Male Broilers Chall - 2017 - PoultrDocument10 pagesEffects of Commercial Organic Acid Blends On Male Broilers Chall - 2017 - PoultrCristian José CardozoNo ratings yet
- Animals 11 01865Document15 pagesAnimals 11 01865Erwin Luis KleinNo ratings yet
- Estudio Mazzota Temperatura PasteurizaciónDocument6 pagesEstudio Mazzota Temperatura PasteurizaciónMitzy CuevasNo ratings yet
- HindawiDocument7 pagesHindawiAsis QimnaaraNo ratings yet
- BMC Vet Res 2013,9-81Document9 pagesBMC Vet Res 2013,9-81Antonia GounadakiNo ratings yet
- Antimicrobial Properties of Salt Natural CasingsDocument6 pagesAntimicrobial Properties of Salt Natural CasingsdesmadradorNo ratings yet
- Carvacrol Empanado de FrangoDocument7 pagesCarvacrol Empanado de FrangoJoel JrNo ratings yet
- Hari Krishnan 2011Document8 pagesHari Krishnan 2011Joshito MoriNo ratings yet
- Biogenic Amine Levels in Dry Fermented Sausages Produced and Sold in GreeceDocument6 pagesBiogenic Amine Levels in Dry Fermented Sausages Produced and Sold in GreeceVeronica PopescuNo ratings yet
- Fate of Spoilage and Pathogenic Microorganisms in Acidified Cold-Filled Hot Pepper SaucesDocument9 pagesFate of Spoilage and Pathogenic Microorganisms in Acidified Cold-Filled Hot Pepper SaucesCaroline MeinertNo ratings yet
- J Sci Food Agric - 1999 - El Alim - Culinary Herbs Inhibit Lipid Oxidation in Raw and Cooked Minced Meat Patties DuringDocument9 pagesJ Sci Food Agric - 1999 - El Alim - Culinary Herbs Inhibit Lipid Oxidation in Raw and Cooked Minced Meat Patties Duringbiancap00No ratings yet
- Intrinsic Parameters QDocument7 pagesIntrinsic Parameters QshelbsNo ratings yet
- Food Control: Leonardo M. Pérez, Marina Del V. Soazo, Claudia E. Balagué, Amelia C. Rubiolo, Roxana A. VerdiniDocument7 pagesFood Control: Leonardo M. Pérez, Marina Del V. Soazo, Claudia E. Balagué, Amelia C. Rubiolo, Roxana A. VerdinirafacpereiraNo ratings yet
- Identification of Staphylococcus Aureus and Escherichia Coli Isolated From Egyptian Food by Conventional and Molecular MethodsDocument7 pagesIdentification of Staphylococcus Aureus and Escherichia Coli Isolated From Egyptian Food by Conventional and Molecular Methodssarim AzizNo ratings yet
- Effects of Boiling Time On The Concentrations of Vitamin C and Betacarotene Infive Selected Green Vegetables Consumed in GhanaDocument6 pagesEffects of Boiling Time On The Concentrations of Vitamin C and Betacarotene Infive Selected Green Vegetables Consumed in GhanaBrandeice BarrettNo ratings yet
- Acidifiers in Animal NutritionDocument1 pageAcidifiers in Animal NutritionJignesh MovaliyaNo ratings yet
- Food Preservation by Hurdle TechnologyDocument7 pagesFood Preservation by Hurdle TechnologyProf C.S.PurushothamanNo ratings yet
- Application of High Pressure Processing To Kill Escherichia Coli O157 in Ready-to-Eat MeatsDocument8 pagesApplication of High Pressure Processing To Kill Escherichia Coli O157 in Ready-to-Eat Meatsgs64569No ratings yet
- HTTP WWW - Sciencedirect.com Science Ob MImg& Imagekey B6T6P-4VJ0CWR-5-7& Cdi 5036& User 687350& Search& CoverDate 06/30/2009& Ie SdarticleDocument5 pagesHTTP WWW - Sciencedirect.com Science Ob MImg& Imagekey B6T6P-4VJ0CWR-5-7& Cdi 5036& User 687350& Search& CoverDate 06/30/2009& Ie SdarticlePaola CastroNo ratings yet
- International Journal of Food Microbiology: E. Apostolidis, Y.-I. Kwon, K. ShettyDocument8 pagesInternational Journal of Food Microbiology: E. Apostolidis, Y.-I. Kwon, K. Shettylordmoises MilanèsNo ratings yet
- Preservation of Blue-Jack Mackerel (Trachurus Picturatus Bowdich) Silage by Chemical and Fermentative AcidificationDocument15 pagesPreservation of Blue-Jack Mackerel (Trachurus Picturatus Bowdich) Silage by Chemical and Fermentative AcidificationMaria DapkeviciusNo ratings yet
- Foods: Influence of Protein Type On The Antimicrobial Activity of LAE Alone or in Combination With MethylparabenDocument14 pagesFoods: Influence of Protein Type On The Antimicrobial Activity of LAE Alone or in Combination With MethylparabenYefii FrisillaNo ratings yet
- The Survival of Escherichia Coli O157:H7 in The Presence of Penicillium Expansum and Glomerella Cingulata in Wounds On Apple SurfacesDocument7 pagesThe Survival of Escherichia Coli O157:H7 in The Presence of Penicillium Expansum and Glomerella Cingulata in Wounds On Apple SurfacesMarwa aljarahNo ratings yet
- Food Chemistry: Elena Peñas, Juana Frias, Beatriz Sidro, Concepción Vidal-ValverdeDocument8 pagesFood Chemistry: Elena Peñas, Juana Frias, Beatriz Sidro, Concepción Vidal-ValverdeKM_147No ratings yet
- Weak Organic Acids in Food Technology: R. Yasothai and R. GiriprasadDocument3 pagesWeak Organic Acids in Food Technology: R. Yasothai and R. GiriprasadFardin XNo ratings yet
- J Scitotenv 2013 06 027Document9 pagesJ Scitotenv 2013 06 027Kumar NeerajNo ratings yet
- Efficacy of Chlorine, Acidic Electrolyzed Water and Aqueous Chlorine Dioxide Solutions To Decontaminate Escherichia Coli O157:H7 From Lettuce LeavesDocument7 pagesEfficacy of Chlorine, Acidic Electrolyzed Water and Aqueous Chlorine Dioxide Solutions To Decontaminate Escherichia Coli O157:H7 From Lettuce LeavesCarolina100% (1)
- Ajas24 12Document8 pagesAjas24 12ahmed galalNo ratings yet
- MicrobiologicalDocument12 pagesMicrobiologicalTrung Truong NhatNo ratings yet
- Listeria in Foods: Collaborative Study: TECRA Listeria Visual Immunoassay (TLVIA) For Detection ofDocument18 pagesListeria in Foods: Collaborative Study: TECRA Listeria Visual Immunoassay (TLVIA) For Detection ofdanzan1No ratings yet
- 2001 Unal Novel Quantitative Analysis Antimicrobial ActivityDocument6 pages2001 Unal Novel Quantitative Analysis Antimicrobial ActivityYeni SulistiyaniNo ratings yet
- Rabbirt SpirullinaDocument5 pagesRabbirt SpirullinaAyvz Martija LambioNo ratings yet
- Non O157 StecDocument4 pagesNon O157 StecEduardo Ramirez LasoNo ratings yet
- 1-S2.0-S0956713516304546-Main Calor EjemploDocument5 pages1-S2.0-S0956713516304546-Main Calor EjemplohergofraNo ratings yet
- Ramaswamy2008 PDFDocument8 pagesRamaswamy2008 PDFnini456No ratings yet
- Red Onion 2Document6 pagesRed Onion 2Mirza RiskNo ratings yet
- Ann. Anim. Sci., Vol. 13, No. 1 (2013) 85-96, DOI: 10.2478/v10220-012-0061-zDocument12 pagesAnn. Anim. Sci., Vol. 13, No. 1 (2013) 85-96, DOI: 10.2478/v10220-012-0061-zMaulana SadidNo ratings yet
- Articulo 3Document5 pagesArticulo 3zayrem montoyaNo ratings yet
- Characterization of E. Coli Strains Obtained From Wastewater EffluentDocument8 pagesCharacterization of E. Coli Strains Obtained From Wastewater EffluentSEP-PublisherNo ratings yet
- Anthocyanin, Phenolics and Antioxidant Activity Changes in Purple Waxy Corn As Affected by Traditional CookingDocument9 pagesAnthocyanin, Phenolics and Antioxidant Activity Changes in Purple Waxy Corn As Affected by Traditional CookingNely Sacsi HuamaniNo ratings yet
- FRAP, DPPH, ABTS and Folin-Ciocalteu MethodsDocument8 pagesFRAP, DPPH, ABTS and Folin-Ciocalteu MethodsAndhika Bintang MahardhikaNo ratings yet
- Food Chemistry: Hua-Bin Li, Ka-Wing Cheng, Chi-Chun Wong, King-Wai Fan, Feng Chen, Yue JiangDocument6 pagesFood Chemistry: Hua-Bin Li, Ka-Wing Cheng, Chi-Chun Wong, King-Wai Fan, Feng Chen, Yue JiangChristopherNo ratings yet
- Protective mechanism of reduced water against alloxan-induced pancreatic β-cell damage: Scavenging effect against reactive oxygen speciesDocument11 pagesProtective mechanism of reduced water against alloxan-induced pancreatic β-cell damage: Scavenging effect against reactive oxygen speciesAditiya Liverpudlian GenkNo ratings yet
- Aeromonas Hydrophila in Biofloc Systems Using Different Carbon SourcesDocument28 pagesAeromonas Hydrophila in Biofloc Systems Using Different Carbon SourcesYousaf JamalNo ratings yet
- Determination of The Degree of Hydrolysis of Food Protein Hydrolysates byDocument8 pagesDetermination of The Degree of Hydrolysis of Food Protein Hydrolysates byHisExcellencyNo ratings yet
- Enteroc y List en Productos SecosDocument34 pagesEnteroc y List en Productos SecosEdson EscamillaNo ratings yet
- Nano Spirulina Dietary Supplementation Augments Growth AO & Immuno Reactions, Digestions & Protection of O. Niloticus Against A. Veronii & Some Physical Stressors, Elabd Et Al., 2020Document13 pagesNano Spirulina Dietary Supplementation Augments Growth AO & Immuno Reactions, Digestions & Protection of O. Niloticus Against A. Veronii & Some Physical Stressors, Elabd Et Al., 2020deryhermawanNo ratings yet
- Factors Affecting Growth and SurvivalDocument26 pagesFactors Affecting Growth and Survivalurmila choudharyNo ratings yet
- Physical-Chemical Properties of Foods: New Tools for PredictionFrom EverandPhysical-Chemical Properties of Foods: New Tools for PredictionNo ratings yet
- Biodegradability of SurfactantsDocument1 pageBiodegradability of SurfactantsTinnysumardiNo ratings yet
- Calibration, Validation and Verification of Equipment, Instruments and Other DevicesDocument2 pagesCalibration, Validation and Verification of Equipment, Instruments and Other DevicesTinnysumardiNo ratings yet
- Minireview Escherichia Coli in The Environment: Implications For Water Quality and Human HealthDocument8 pagesMinireview Escherichia Coli in The Environment: Implications For Water Quality and Human HealthTinnysumardiNo ratings yet
- Zoning in Processing Facilities 375003 7Document2 pagesZoning in Processing Facilities 375003 7TinnysumardiNo ratings yet
- Federal Register / Vol. 60, No. 183 / Thursday, September 21, 1995 / Proposed RulesDocument2 pagesFederal Register / Vol. 60, No. 183 / Thursday, September 21, 1995 / Proposed RulesTinnysumardiNo ratings yet
- Escherichia Coli O157:H7 Issues and Ramifications: Executive SummaryDocument12 pagesEscherichia Coli O157:H7 Issues and Ramifications: Executive SummaryTinnysumardiNo ratings yet
- Rlu PDFDocument7 pagesRlu PDFTinnysumardiNo ratings yet
- Monitoring Suhu Dan RH WH Starch 2Document4 pagesMonitoring Suhu Dan RH WH Starch 2TinnysumardiNo ratings yet
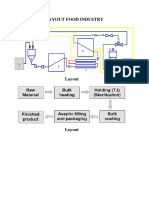
- Layout Food IndustryDocument2 pagesLayout Food IndustryTinnysumardiNo ratings yet
- BookDocument155 pagesBookTinnysumardiNo ratings yet
- Infant Formula RecallDocument4 pagesInfant Formula RecallTinnysumardiNo ratings yet
- International Formula Council: Formerly The Enteral Nutrition Council and Infant Formula CouncilDocument6 pagesInternational Formula Council: Formerly The Enteral Nutrition Council and Infant Formula CouncilTinnysumardiNo ratings yet
- Science NotesDocument10 pagesScience NotesLiezel BrillantesNo ratings yet
- Normal Hemodynamic Parameters and Lab Values - Reference CardDocument4 pagesNormal Hemodynamic Parameters and Lab Values - Reference Cardblanquishem100% (1)
- CARBOHYDRATES WPS OfficeDocument11 pagesCARBOHYDRATES WPS OfficeSyainaaa DalpasanNo ratings yet
- 4-D Printing Technology: Department of Mechanical Engineering K.V.G. College of Engineering Sullia, (D.K.) 574327Document12 pages4-D Printing Technology: Department of Mechanical Engineering K.V.G. College of Engineering Sullia, (D.K.) 574327JECINJOHNNo ratings yet
- Res Ifu TPH Spectra ST Effects 2020 05Document56 pagesRes Ifu TPH Spectra ST Effects 2020 05Karen BeleñoNo ratings yet
- Daftar Pustaka: Endomycorrhizas. Academic Press, London. In: Mawarni, S.A. 2011Document9 pagesDaftar Pustaka: Endomycorrhizas. Academic Press, London. In: Mawarni, S.A. 2011Muh Nur TegarNo ratings yet
- Is Aloe Vera Effective For Wound Healing? The State of The ArtDocument6 pagesIs Aloe Vera Effective For Wound Healing? The State of The Artwulan kusuma wardaniNo ratings yet
- Pierluigi IghinaDocument38 pagesPierluigi Ighinaemmepicsd100% (2)
- Forensic Dentistry/Odontology: James HerreraDocument13 pagesForensic Dentistry/Odontology: James HerrerackamilkywayNo ratings yet
- Intro To Stat (STAT 111) by EwensDocument113 pagesIntro To Stat (STAT 111) by EwensIris ZhangNo ratings yet
- Breeding Seasons and Lambing SeasonsDocument16 pagesBreeding Seasons and Lambing SeasonsVijaya lakshmiNo ratings yet
- Basidiomycota: Mushrooms, Bracket Fungi, Puffballs, Rusts and SmutsDocument76 pagesBasidiomycota: Mushrooms, Bracket Fungi, Puffballs, Rusts and SmutsaaronNo ratings yet
- Lista Cartilor Elsevier AchizitionateDocument57 pagesLista Cartilor Elsevier AchizitionateLucica PleșcaNo ratings yet
- Introduction To AnthropologyDocument17 pagesIntroduction To AnthropologyJaycee TualaNo ratings yet
- Miyamoto 2010 JBJS Current Concepts Review Distal-Biceps-Tendon-Injuries PDFDocument12 pagesMiyamoto 2010 JBJS Current Concepts Review Distal-Biceps-Tendon-Injuries PDFKuozram No MasNo ratings yet
- Iron and Sulfur Oxidizing BacteriaDocument11 pagesIron and Sulfur Oxidizing BacterianadyatulhanifNo ratings yet
- Application of Artificial Intelligence in Medical FieldDocument8 pagesApplication of Artificial Intelligence in Medical FieldShree krishana ShresthaNo ratings yet
- Different Operating Room InstrumentDocument4 pagesDifferent Operating Room InstrumentMaikka Kyril Abarquez67% (3)
- 5991 1876enDocument32 pages5991 1876enBeatriz ReyesNo ratings yet
- Axel Zarske Systematik Einiger Blutsalmler Oder Rosy TetrasDocument30 pagesAxel Zarske Systematik Einiger Blutsalmler Oder Rosy Tetraskatral1968No ratings yet
- 1-20 Mock Tests CATDocument872 pages1-20 Mock Tests CATTanmay GargNo ratings yet
- 30 of BodybuildingDocument10 pages30 of Bodybuildingariven11No ratings yet
- Sample: Body Fluid Analysis For Cellular Composition Approved GuidelineDocument13 pagesSample: Body Fluid Analysis For Cellular Composition Approved GuidelineBryan Guisberth RojasNo ratings yet
- Basic of BiochemistryDocument3 pagesBasic of BiochemistrySandy AHNo ratings yet
- Gods and Monsters PDFDocument62 pagesGods and Monsters PDFDiego Muniz100% (2)
- 00 - Contents Everyday Science MCQsDocument5 pages00 - Contents Everyday Science MCQsmimtiazshahid100% (1)
- Cancer Cure Booklet - Drkelleyenzymes PDFDocument21 pagesCancer Cure Booklet - Drkelleyenzymes PDFSengNo ratings yet