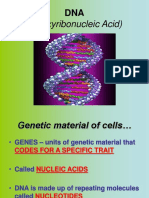
You might also like
- Hybridization with Nucleic Acid Probes, Part II: Part II. Probe Labeling and Hybridization TechniquesFrom EverandHybridization with Nucleic Acid Probes, Part II: Part II. Probe Labeling and Hybridization TechniquesNo ratings yet
- Chromosome Structure and FunctionDocument76 pagesChromosome Structure and Functionstevensb055No ratings yet
- LET'S MAKE NEW CELLSDocument10 pagesLET'S MAKE NEW CELLSDaniel BerryNo ratings yet
- Lecture 2 (DNA Replication)Document37 pagesLecture 2 (DNA Replication)M Luqman Hakim100% (1)
- Basics of Molecular BiologyDocument69 pagesBasics of Molecular BiologypathinfoNo ratings yet
- Chromosomes: Sardar Hussain Asst - Prof.biotechnology GSC, CtaDocument127 pagesChromosomes: Sardar Hussain Asst - Prof.biotechnology GSC, CtaAshikuzzaman AntorNo ratings yet
- Gel Electrophoresis PDFDocument15 pagesGel Electrophoresis PDFTeflon Slim100% (1)
- MeiosisDocument24 pagesMeiosisMuhammad HamzaNo ratings yet
- Gene RegulationDocument30 pagesGene Regulationprehealthhelp67% (3)
- Chromosomes: The Blueprint of LifeDocument16 pagesChromosomes: The Blueprint of LifeJan Lloyd OdruniaNo ratings yet
- DNA and Protein Synthesis - Chapter Notes: "Life Is A Three Letter Word!"Document13 pagesDNA and Protein Synthesis - Chapter Notes: "Life Is A Three Letter Word!"Chan Karlok100% (1)
- Genetics, Chapter 3, DNA Replication Lectures (Slides)Document115 pagesGenetics, Chapter 3, DNA Replication Lectures (Slides)Ali Al-QudsiNo ratings yet
- Biotech Principles: Genetic Engineering & Microbial GrowthDocument6 pagesBiotech Principles: Genetic Engineering & Microbial GrowthIndrajith MjNo ratings yet
- Restriction EnzymesDocument14 pagesRestriction EnzymesKnowinter0% (2)
- Central Dogma of Molecular BiologyDocument6 pagesCentral Dogma of Molecular BiologyKim GojoCruzNo ratings yet
- Gel ElectrophoresisDocument6 pagesGel Electrophoresisapi-385481351No ratings yet
- Lecture 1 - Chromosome & GenomeDocument64 pagesLecture 1 - Chromosome & GenomeFriendlyGoodGirlNo ratings yet
- DNA Structure and ReplicationDocument48 pagesDNA Structure and Replicationfatzzahra100% (1)
- Edexcel GCE Biology Unit 2 Exam Revision NotesDocument18 pagesEdexcel GCE Biology Unit 2 Exam Revision NotescolachauNo ratings yet
- Genetic Control of Protein Structure & FunctionDocument60 pagesGenetic Control of Protein Structure & Functionathirah100% (1)
- Regulation of Gene ExpressionDocument20 pagesRegulation of Gene ExpressionrainabtNo ratings yet
- Study Guide - Gene Regulation (CH 16)Document5 pagesStudy Guide - Gene Regulation (CH 16)jaysusgagaNo ratings yet
- DNA Manipulative EnzymesDocument17 pagesDNA Manipulative EnzymesZain Ul AbedienNo ratings yet
- Classical Mendelian GeneticsDocument33 pagesClassical Mendelian GeneticsAngel Pearl DeneroNo ratings yet
- Cell Cycle Review KeyDocument4 pagesCell Cycle Review KeyaikaterinaNo ratings yet
- Transgenic Animals: Their Benefits To Human Welfare: What Is A Transgenic Animal?Document7 pagesTransgenic Animals: Their Benefits To Human Welfare: What Is A Transgenic Animal?anon_939844203No ratings yet
- Characteristics and Genotyping (Semi-Automated and Automated), Apparatus Used in GenotypingDocument45 pagesCharacteristics and Genotyping (Semi-Automated and Automated), Apparatus Used in GenotypingKhalid HameedNo ratings yet
- Cell - The Unit of LifeDocument71 pagesCell - The Unit of LifeGanesh PatilNo ratings yet
- Cell TheoryDocument10 pagesCell TheoryLuigi Miguel G. TiranaNo ratings yet
- DNA ReplicationDocument83 pagesDNA ReplicationVipin100% (8)
- Mitosis and MeiosisDocument56 pagesMitosis and MeiosisJaime Soriano100% (1)
- Practice Questions For Dna Replication and RepairDocument11 pagesPractice Questions For Dna Replication and RepairdhruviniNo ratings yet
- cDNA Libraries and Gene CloningDocument8 pagescDNA Libraries and Gene CloningRoberto RomeroNo ratings yet
- Linkage: Harshraj Subhash Shinde KKW, Cabt, NashikDocument14 pagesLinkage: Harshraj Subhash Shinde KKW, Cabt, Nashiksivaram888No ratings yet
- Answering Exam Questions On CellsDocument4 pagesAnswering Exam Questions On CellsTamicka Bonnick100% (1)
- Cell CycleDocument29 pagesCell Cycleolowolo mutiuNo ratings yet
- AP Bio CH 18-20 Take Home FinalDocument14 pagesAP Bio CH 18-20 Take Home Finalheeraangelwingz0% (1)
- Protein Synthesis Transcription and Translation Distance LearningDocument10 pagesProtein Synthesis Transcription and Translation Distance LearningjaneyzhouNo ratings yet
- Deoxyribonucleic AcidDocument36 pagesDeoxyribonucleic AcidAlex LlanoNo ratings yet
- DNA Structure & ReplicationDocument25 pagesDNA Structure & Replicationivanam0202No ratings yet
- CH - 40 - Introduction To The Cell CycleDocument15 pagesCH - 40 - Introduction To The Cell Cycleerichaas100% (1)
- 1.0 Genetics and InheritanceDocument13 pages1.0 Genetics and Inheritanceincognitus94100% (3)
- Protein Synthesis - Transcription and TranslationDocument61 pagesProtein Synthesis - Transcription and Translationblackmoneygrabber100% (1)
- 11.1 Antibody Production and VaccinationDocument28 pages11.1 Antibody Production and VaccinationFRENCHONLYNo ratings yet
- Plant vs Animal Cell DifferencesDocument6 pagesPlant vs Animal Cell DifferencesYukiNo ratings yet
- Intro to Bio Diversity - Taxonomy, Classification & PhylogenyDocument3 pagesIntro to Bio Diversity - Taxonomy, Classification & PhylogenyNurul Amirah HossainNo ratings yet
- Genes on ChromosomesDocument31 pagesGenes on Chromosomespraata123No ratings yet
- DNA Structure and Function - First Lecture (4822)Document17 pagesDNA Structure and Function - First Lecture (4822)Ashley Ramos LópezNo ratings yet
- Mendelian & Non-Mendelian GeneticsDocument37 pagesMendelian & Non-Mendelian GeneticsJubair Al-rashid100% (2)
- Cell and Molecular Biology Module (Lecture and Laboratory)Document200 pagesCell and Molecular Biology Module (Lecture and Laboratory)RM Montemayor100% (2)
- Cell Structure and FunctionDocument41 pagesCell Structure and FunctionAhkyluzLaniazNo ratings yet
- Meiosis Explained: The Process That Halves Chromosome NumberDocument38 pagesMeiosis Explained: The Process That Halves Chromosome NumberkrystenNo ratings yet
- Crispr Cas HajarDocument21 pagesCrispr Cas HajarHajira Fatima100% (1)
- DNA Replication in ProkaryotesDocument24 pagesDNA Replication in ProkaryotesAlbert Jade Pontimayor Legaria100% (1)
- t2 Bio Revision Ex 1 Answer SchemeDocument15 pagest2 Bio Revision Ex 1 Answer SchemeNicholas OwNo ratings yet
- Ch10 Mendelian GeneticsDocument42 pagesCh10 Mendelian GeneticseubacteriaNo ratings yet
- Directed Mutagenesis and Protein EngineeringDocument52 pagesDirected Mutagenesis and Protein Engineeringslowdragon2003No ratings yet
- Understanding ChromosomesDocument40 pagesUnderstanding ChromosomesinezNo ratings yet
- Module 2 - Cellular Basis of InheritanceDocument148 pagesModule 2 - Cellular Basis of InheritanceJeremy GyamfiNo ratings yet
- Eukaryotic ChromosomesDocument8 pagesEukaryotic Chromosomesfariha munawwerNo ratings yet
- Universiti Malaysia Sabah Faculty of Science and Human Resources Hgo7 BiotechnologyDocument9 pagesUniversiti Malaysia Sabah Faculty of Science and Human Resources Hgo7 BiotechnologyShalini MuthuNo ratings yet
- Genetics Molecules AssignmentDocument4 pagesGenetics Molecules AssignmentShalini MuthuNo ratings yet
- MBA711 - Answers To All Chapter 7 ProblemsDocument21 pagesMBA711 - Answers To All Chapter 7 Problemsshweta shuklaNo ratings yet
- Length of Roots and Shoot Vs Sucrose TreatmentDocument2 pagesLength of Roots and Shoot Vs Sucrose TreatmentShalini MuthuNo ratings yet
- Malaysian Coffee - Coffee of Malaysia 3 May 2018. Retrieved FromDocument2 pagesMalaysian Coffee - Coffee of Malaysia 3 May 2018. Retrieved FromShalini MuthuNo ratings yet
- ExerciseDocument2 pagesExerciseShalini MuthuNo ratings yet
- Diagram of MonopolyDocument3 pagesDiagram of MonopolyShalini MuthuNo ratings yet
- Cappuccino GDocument1 pageCappuccino GShalini MuthuNo ratings yet
- Physics Group AssignmentDocument10 pagesPhysics Group AssignmentShalini MuthuNo ratings yet
- MueteassyDocument4 pagesMueteassyShalini MuthuNo ratings yet
- The history and science behind the iconic appleDocument5 pagesThe history and science behind the iconic appleShalini MuthuNo ratings yet
- The history and science behind the iconic appleDocument5 pagesThe history and science behind the iconic appleShalini MuthuNo ratings yet
- Learning Online: Veletsianos, GeorgeDocument11 pagesLearning Online: Veletsianos, GeorgePsico XavierNo ratings yet
- Delhi Public School: Class: XI Subject: Assignment No. 3Document1 pageDelhi Public School: Class: XI Subject: Assignment No. 3Aman Kumar BhagatNo ratings yet
- Donny UfoaksesDocument27 pagesDonny UfoaksesKang Bowo D'wizardNo ratings yet
- BMXNRPDocument60 pagesBMXNRPSivaprasad KcNo ratings yet
- Trimble Oem Gnss Bro Usl 0422Document3 pagesTrimble Oem Gnss Bro Usl 0422rafaelNo ratings yet
- Ipo Exam Revised SyllabusDocument1 pageIpo Exam Revised Syllabusজ্যোতিৰ্ময় বসুমতাৰীNo ratings yet
- RestrukturisasiDocument17 pagesRestrukturisasimuhammad hayel wallaNo ratings yet
- Av1 OnDocument7 pagesAv1 OnLê Hà Thanh TrúcNo ratings yet
- Useful Coaching Questions: Questions To Create A State Change Questions To Ask When Something Goes WrongDocument2 pagesUseful Coaching Questions: Questions To Create A State Change Questions To Ask When Something Goes WrongAntonioNo ratings yet
- E PortfolioDocument76 pagesE PortfolioMAGALLON ANDREWNo ratings yet
- Product Catalog 2016Document84 pagesProduct Catalog 2016Sauro GordiniNo ratings yet
- Hotel Design Planning and DevelopmentDocument30 pagesHotel Design Planning and DevelopmentTio Yogatma Yudha14% (7)
- EMECH 2 MarksDocument18 pagesEMECH 2 MarkspavanraneNo ratings yet
- EE-434 Power Electronics: Engr. Dr. Hadeed Ahmed SherDocument23 pagesEE-434 Power Electronics: Engr. Dr. Hadeed Ahmed SherMirza Azhar HaseebNo ratings yet
- LLM DissertationDocument94 pagesLLM Dissertationjasminjajarefe100% (1)
- Main Hoon Na - WikipediaDocument8 pagesMain Hoon Na - WikipediaHusain ChandNo ratings yet
- WA Beretta M92FS Parts ListDocument2 pagesWA Beretta M92FS Parts ListDenis Deki NehezNo ratings yet
- Accidental PoisoningDocument3 pagesAccidental PoisoningBRUELIN MELSHIA MNo ratings yet
- PM - Network Analysis CasesDocument20 pagesPM - Network Analysis CasesImransk401No ratings yet
- Plant Air Centrifugal Compressors: Turbo-Air Series Featuring Oil-Free AirDocument20 pagesPlant Air Centrifugal Compressors: Turbo-Air Series Featuring Oil-Free AirSharad KokateNo ratings yet
- Fisher FIELDVUE DVC2000 Digital Valve Controller: Instruction ManualDocument108 pagesFisher FIELDVUE DVC2000 Digital Valve Controller: Instruction ManualsrinuvoodiNo ratings yet
- COT EnglishDocument4 pagesCOT EnglishTypie ZapNo ratings yet
- Philippine College of Northwestern Luzon Bachelor of Science in Business AdministrationDocument7 pagesPhilippine College of Northwestern Luzon Bachelor of Science in Business Administrationzackwayne100% (1)
- Brooks Instrument FlowmeterDocument8 pagesBrooks Instrument FlowmeterRicardo VillalongaNo ratings yet
- Revised Man As A Biological BeingDocument8 pagesRevised Man As A Biological Beingapi-3832208No ratings yet
- Intec Waste PresiDocument8 pagesIntec Waste Presiapi-369931794No ratings yet
- Complete Hemi Sync Gateway Experience ManualDocument43 pagesComplete Hemi Sync Gateway Experience Manualapi-385433292% (92)
- Addition and Subtraction of PolynomialsDocument8 pagesAddition and Subtraction of PolynomialsPearl AdamosNo ratings yet
- Week 6Document7 pagesWeek 6Nguyễn HoàngNo ratings yet