You might also like
- 2017 - Re-Understanding Anti-Influenza Strategy Attach Equal Importance To Antiviral and Anti-Inflammatory TherapiesDocument12 pages2017 - Re-Understanding Anti-Influenza Strategy Attach Equal Importance To Antiviral and Anti-Inflammatory TherapiesVladDaculNo ratings yet
- 2011 - Metformin - Multi-Faceted Protection Against CancerDocument22 pages2011 - Metformin - Multi-Faceted Protection Against CancerVladDaculNo ratings yet
- 2013 - Reductive Glutamine Metabolism by IDH1 Mediates Lipogenesis Under HypoxiaDocument10 pages2013 - Reductive Glutamine Metabolism by IDH1 Mediates Lipogenesis Under HypoxiaVladDaculNo ratings yet
- Living Water - Braun & Cavagnaro 2Document9 pagesLiving Water - Braun & Cavagnaro 2VladDaculNo ratings yet
- (The-Persecution-and-Trial-of-Gaston-Naessens) OK PDFDocument221 pages(The-Persecution-and-Trial-of-Gaston-Naessens) OK PDFVladDacul100% (1)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Apoptosis UsmleDocument2 pagesApoptosis Usmlelovelyc95100% (1)
- Quiz 6Document4 pagesQuiz 6JeevikaGoyal100% (1)
- Lecture 3 - Enzyme and Enzyme Kinetics PDFDocument8 pagesLecture 3 - Enzyme and Enzyme Kinetics PDFJulius BersabeNo ratings yet
- GLIKOGENESISDocument45 pagesGLIKOGENESISChrista24796No ratings yet
- Paper I MCQ's PhysiologyDocument32 pagesPaper I MCQ's PhysiologyMadhu RauniyarNo ratings yet
- June 2019 (IAL) MS - Unit 1 Edexcel Biology A-LevelDocument28 pagesJune 2019 (IAL) MS - Unit 1 Edexcel Biology A-LevelAmmar ZaffarullahNo ratings yet
- 6Document17 pages6AmaniNo ratings yet
- Structural Analysis of Sequence Data in Virology An Elementary Approach Using Sars-Cov-2 As An ExampleDocument35 pagesStructural Analysis of Sequence Data in Virology An Elementary Approach Using Sars-Cov-2 As An ExampleAdrian BoboceaNo ratings yet
- ProteinDocument32 pagesProteinYo1No ratings yet
- MIMM 211 MidtermDocument8 pagesMIMM 211 MidtermIpshitanandi100% (1)
- Biology Class 10 Icse Boards!Document10 pagesBiology Class 10 Icse Boards!nark.jf4No ratings yet
- Proteins 3: 2°& 3°structure & F Ldi D ST Bilit Folding and StabilityDocument34 pagesProteins 3: 2°& 3°structure & F Ldi D ST Bilit Folding and StabilityshaheenNo ratings yet
- Nervous Systems - Neuron Function: Excitability Resting Potentials Graded Potential Action Potential The SynapseDocument48 pagesNervous Systems - Neuron Function: Excitability Resting Potentials Graded Potential Action Potential The SynapseRajeev AgarwalNo ratings yet
- Bio Project FileDocument12 pagesBio Project FileharshitNo ratings yet
- Biochemistry For Psychiatry Students by Abayneh EDocument123 pagesBiochemistry For Psychiatry Students by Abayneh Egobez temariNo ratings yet
- Experiment 2 - Enzyme ActivityDocument7 pagesExperiment 2 - Enzyme ActivityFatimatuzzahra' Binti Hardiyono FISNo ratings yet
- 02.06.2018 - 5 - M. Vives Santacana - Principles of PharmacodynamicsDocument46 pages02.06.2018 - 5 - M. Vives Santacana - Principles of PharmacodynamicsPhilippe KinnaerNo ratings yet
- Enzyme Kinetics ProblemsDocument5 pagesEnzyme Kinetics ProblemsBlessy Gabayno100% (1)
- Section 8-2 Notes 2022-2023Document12 pagesSection 8-2 Notes 2022-2023Verificar Ameijenda PratoNo ratings yet
- Ichthyosis: A Road Model For Skin Research: Review ArticleDocument10 pagesIchthyosis: A Road Model For Skin Research: Review ArticlebaihaqiNo ratings yet
- CellStructureSEDocument3 pagesCellStructureSEJin KingNo ratings yet
- β-Thalassemia: ReviewDocument11 pagesβ-Thalassemia: ReviewSajjad Hossain ShuvoNo ratings yet
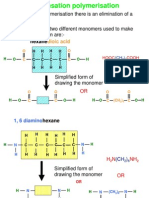
- Condensation PolymerisationDocument14 pagesCondensation Polymerisationafoo1234100% (2)
- Genetic Control of ProteinDocument43 pagesGenetic Control of ProteinGabriela ZahiuNo ratings yet
- The Aspects of Gene TherapyDocument9 pagesThe Aspects of Gene TherapyCharice Anne VillamarinNo ratings yet
- B.SC Biotech Syllabus-1Document41 pagesB.SC Biotech Syllabus-1Dr. Maruti K. R SDM Degree College UjireNo ratings yet
- Unit 1: The Cell 5Th GradeDocument2 pagesUnit 1: The Cell 5Th GradeAriadna AtencioNo ratings yet
- SBI-4U Unit 1 TestDocument6 pagesSBI-4U Unit 1 TestpokesurferNo ratings yet
- Microbiology: Basic and Clinical Principles: First EditionDocument112 pagesMicrobiology: Basic and Clinical Principles: First EditionDawn DixonNo ratings yet
- OrabyBiochemistry P.2 PDFDocument232 pagesOrabyBiochemistry P.2 PDFJayadev Ch100% (1)