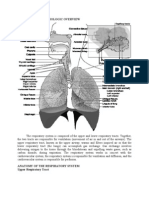
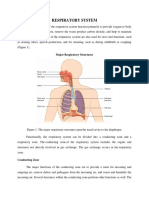
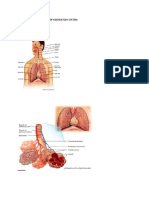
You might also like
- Treatise on the Anatomy and Physiology of the Mucous Membranes: With Illustrative Pathological ObservationsFrom EverandTreatise on the Anatomy and Physiology of the Mucous Membranes: With Illustrative Pathological ObservationsNo ratings yet
- Histology of Larynx, Trachea, Bronchus - Dea Natalia - 130110110190 - E3/RespiDocument3 pagesHistology of Larynx, Trachea, Bronchus - Dea Natalia - 130110110190 - E3/RespiMarshel BudiarsaNo ratings yet
- Animal CellDocument64 pagesAnimal CellFrance Dorothy SamortinNo ratings yet
- Respiratory Epithelium: Human Histology Lecture Chapter 17 Ï The Respiratory SystemDocument16 pagesRespiratory Epithelium: Human Histology Lecture Chapter 17 Ï The Respiratory SystemEvandie OngNo ratings yet
- Respiratory System 2Document8 pagesRespiratory System 2Shawn Gaurav JhaNo ratings yet
- 1st Lecture of Respiratory Histology by DR RoomiDocument24 pages1st Lecture of Respiratory Histology by DR RoomiMudassar Roomi100% (1)
- The Respiratory System PP First - Stage 2021Document27 pagesThe Respiratory System PP First - Stage 2021HHHNo ratings yet
- (Histologi) Sistem Respirasi - Dr. SusiantiDocument51 pages(Histologi) Sistem Respirasi - Dr. SusiantiulfamarianaNo ratings yet
- Respiratory System Anatomy and FunctionDocument12 pagesRespiratory System Anatomy and Functionمحمد العراقيNo ratings yet
- Respiratory System: A) The Conducting Portion, Which Consists of The Nasal CavitiesDocument7 pagesRespiratory System: A) The Conducting Portion, Which Consists of The Nasal CavitiesSalmah Alimah FitriNo ratings yet
- Respiratory SystemDocument25 pagesRespiratory Systemada.codreanuNo ratings yet
- SasuntqiDocument55 pagesSasuntqiNika MtsituriNo ratings yet
- Structure and Function of The Respiratory System: Lung AnatomyDocument1 pageStructure and Function of The Respiratory System: Lung AnatomygotgotaoNo ratings yet
- Histology of Respiratory System: Dr. Mirna Muis, SP - Rad Dept. of Histology Faculty of Medicine, Hasanuddin UnivDocument31 pagesHistology of Respiratory System: Dr. Mirna Muis, SP - Rad Dept. of Histology Faculty of Medicine, Hasanuddin UnivNor Izhharuddin ZainyNo ratings yet
- Respiratory SystemDocument15 pagesRespiratory SystemJovelou MihangosNo ratings yet
- 3 Physiology of The Nose and Paranasal Sinuses: 3.2 BreathingDocument6 pages3 Physiology of The Nose and Paranasal Sinuses: 3.2 BreathingMansour HazaNo ratings yet
- Lecture 12 Respiratory System SHF 2023Document11 pagesLecture 12 Respiratory System SHF 2023Kumru Kaleli Kumru KaleliNo ratings yet
- Respiratory System by Dr. Vincent-JrDocument48 pagesRespiratory System by Dr. Vincent-JrEmma OsabohienNo ratings yet
- Gas Exchange and the Dangers of SmokingDocument15 pagesGas Exchange and the Dangers of SmokingZahina AmrinNo ratings yet
- Ma Nurfaizah Ma04200026 Respiratory System April 2020Document3 pagesMa Nurfaizah Ma04200026 Respiratory System April 2020Nur Faizah Binti Jamail MANo ratings yet
- Dulutalias - Chapter23-THE RESPIRATORY SYSTEMDocument15 pagesDulutalias - Chapter23-THE RESPIRATORY SYSTEMKie MusstisqueNo ratings yet
- AaDocument324 pagesAaBirhanu AyenewNo ratings yet
- Sistem RespirasiDocument27 pagesSistem RespirasiFitri FirifiNo ratings yet
- Functions of The Respiratory SystemDocument98 pagesFunctions of The Respiratory SystemNimra TariqNo ratings yet
- Nasal Cavity and Larynx AnatomyDocument3 pagesNasal Cavity and Larynx AnatomyabdeljelileNo ratings yet
- AICE Bio Notes Pages 186-190: Gas ExchangeDocument2 pagesAICE Bio Notes Pages 186-190: Gas ExchangeSanzida HossainNo ratings yet
- 2nd Lec of Respiratory Histology by DR RoomiDocument39 pages2nd Lec of Respiratory Histology by DR RoomiMudassar Roomi100% (1)
- Anatomy and Physiology of Respiratory SystemDocument4 pagesAnatomy and Physiology of Respiratory Systembilliam123No ratings yet
- Bilology XII 2014 CH - 14 RespirationDocument16 pagesBilology XII 2014 CH - 14 RespirationMuhammad UsmanNo ratings yet
- The Respirator Y System: Mungcal, Dharlynette RTRPDocument158 pagesThe Respirator Y System: Mungcal, Dharlynette RTRPDharlyn MungcalNo ratings yet
- Anatomy and Physiology of The Respiratory SystemDocument57 pagesAnatomy and Physiology of The Respiratory SystemRenz Ivan FuntilonNo ratings yet
- Lect 3Document8 pagesLect 3علي الاسديNo ratings yet
- Anatomic and Physiologic OverviewDocument8 pagesAnatomic and Physiologic OverviewJoseph King MacaranasNo ratings yet
- Respiratory Physiology Part 1Document9 pagesRespiratory Physiology Part 1kabir musa ladanNo ratings yet
- Nasopharynx, Larynx, Trachea, Bronchi, Bronchioles and Terminal BronchiolesDocument3 pagesNasopharynx, Larynx, Trachea, Bronchi, Bronchioles and Terminal BronchiolesMichael UrrutiaNo ratings yet
- Supplementary Material 3.1 Basic Concepts and Assessment (N) - 2Document13 pagesSupplementary Material 3.1 Basic Concepts and Assessment (N) - 2Kent Borro CantomayorNo ratings yet
- Diagnostic Examination of Pulmonary TuberculosisDocument5 pagesDiagnostic Examination of Pulmonary TuberculosisMark BerbanoNo ratings yet
- Normal Histology of The Lung and Pathophysiology of COPD (T Year)Document36 pagesNormal Histology of The Lung and Pathophysiology of COPD (T Year)khaledalameddineNo ratings yet
- Dr Abdul Rehman's Guide to the Respiratory SystemDocument33 pagesDr Abdul Rehman's Guide to the Respiratory SystemMaham BaqaiNo ratings yet
- Histology of Respiratory SystemDocument6 pagesHistology of Respiratory SystemEzyan Syamin100% (1)
- Human Biology: Respiratory SystemDocument33 pagesHuman Biology: Respiratory SystemSaikrishna RaoNo ratings yet
- A & P RespiratoryDocument18 pagesA & P Respiratoryamirul haakimNo ratings yet
- Learning Objective: Respiratory SystemDocument4 pagesLearning Objective: Respiratory SystemReham QueNo ratings yet
- Chapter - 15 The Nose and The Paranasal SinusesDocument6 pagesChapter - 15 The Nose and The Paranasal Sinusesmanojchouhan2014No ratings yet
- Respiratory System A. BDocument5 pagesRespiratory System A. BPhoebe Kates Dela CruzNo ratings yet
- 07 (1) Pulmonary Effects of InhaledDocument46 pages07 (1) Pulmonary Effects of InhaledLeticia MeloNo ratings yet
- Histology of LungsDocument30 pagesHistology of LungsShahir21No ratings yet
- Respiratory System HandoutDocument15 pagesRespiratory System HandoutJei SanNo ratings yet
- Respiratory SystemDocument20 pagesRespiratory SystemKunal DangatNo ratings yet
- Rts k3 Rts1k3Document55 pagesRts k3 Rts1k3felixNo ratings yet
- Pulmo Management Week 1Document160 pagesPulmo Management Week 1Dharlyn MungcalNo ratings yet
- 05 - Chapter 1Document39 pages05 - Chapter 1hamam salih badriNo ratings yet
- Anatomy and Physiology of Respiratory SystemDocument5 pagesAnatomy and Physiology of Respiratory SystemNesra de PeraltaNo ratings yet
- Problems With OxygenationDocument92 pagesProblems With OxygenationEbiNo ratings yet
- 50 The Gas Exchange SystemDocument5 pages50 The Gas Exchange SystemSarahNo ratings yet
- Histology The Respiratory System: Prof. Dr. Sabah N. AlwachiDocument15 pagesHistology The Respiratory System: Prof. Dr. Sabah N. AlwachilaithjnbNo ratings yet
- Pain, Surgery and Oxygenation NursingDocument7 pagesPain, Surgery and Oxygenation NursingJhoanna de ChavezNo ratings yet
- Respiratory System s2 2019Document56 pagesRespiratory System s2 2019noviantyramadhani12No ratings yet
- Respiratory DisordersDocument20 pagesRespiratory DisordersjeromeNo ratings yet
- C612Document5 pagesC612dinhtung2210100% (1)
- Earth First! Climbers Guild Vol. 2Document16 pagesEarth First! Climbers Guild Vol. 2CascadiaDefenders100% (2)
- Polythionic Acid Stress Corrosion CrackingDocument2 pagesPolythionic Acid Stress Corrosion CrackingVajid MadathilNo ratings yet
- NBME 22 OfflineDocument200 pagesNBME 22 OfflineGautham Kanagala86% (14)
- WorkbookEdition 12 A Theory of HumorismDocument4 pagesWorkbookEdition 12 A Theory of HumorismMabel Ferr50% (2)
- Tutorial Sheet 4Document2 pagesTutorial Sheet 4Syed YousufuddinNo ratings yet
- The Prevention of Women & Children Repression Act 2000: A Study of Implementation Process From 2003 To 2013Document9 pagesThe Prevention of Women & Children Repression Act 2000: A Study of Implementation Process From 2003 To 2013Md. Abdul KarimNo ratings yet
- Wheat Sourdough Fermentation - Effects of Time and Acidification On Fundamental Rheological PropertiesDocument9 pagesWheat Sourdough Fermentation - Effects of Time and Acidification On Fundamental Rheological PropertiesmaurodiloretoNo ratings yet
- Inside Out or Outside In. Meeting With Couples. Hugh JenkinsDocument24 pagesInside Out or Outside In. Meeting With Couples. Hugh JenkinsRita1976No ratings yet
- 1978 - 90 Day Study or Rhesus Monkey ToxicityDocument4 pages1978 - 90 Day Study or Rhesus Monkey Toxicitygrace poudrierNo ratings yet
- Indian Pharmacopiea 2007Document73 pagesIndian Pharmacopiea 2007Deepak ShahNo ratings yet
- Come Back To Your Senses Use Your Body: Psychologyt LsDocument1 pageCome Back To Your Senses Use Your Body: Psychologyt LsMarina Moran100% (1)
- Department of The Navy Core Values CharterDocument1 pageDepartment of The Navy Core Values CharterShaun JohnsonNo ratings yet
- ODN102001 ATN Products Hardware Introduction ISSUE 1.00Document43 pagesODN102001 ATN Products Hardware Introduction ISSUE 1.00Jonathan Eduardo Tapias BeltranNo ratings yet
- Corticosteroid: Saut Samuel Simamora Department Ophthalmology Diponegoro University-Kariadi HospitalDocument21 pagesCorticosteroid: Saut Samuel Simamora Department Ophthalmology Diponegoro University-Kariadi HospitalSania NadianisaNo ratings yet
- Derivatives and Risk ManagementDocument17 pagesDerivatives and Risk ManagementDeepak guptaNo ratings yet
- A4 RelationshipgridDocument32 pagesA4 RelationshipgridjamesNo ratings yet
- Different Surgical Modalities For Management of Postburn FL Exion Contracture of The ElbowDocument6 pagesDifferent Surgical Modalities For Management of Postburn FL Exion Contracture of The ElbowMadhuchandra HirehalliNo ratings yet
- Comparing Food with Comparative AdjectivesDocument4 pagesComparing Food with Comparative AdjectivesLuisito GonzalezNo ratings yet
- Psychedelic Drugs-A New Era in Psychiatry?Document9 pagesPsychedelic Drugs-A New Era in Psychiatry?PeterNo ratings yet
- Quiz Integumentary SystemDocument23 pagesQuiz Integumentary SystemLemuel CunananNo ratings yet
- IED Recognition GuideDocument28 pagesIED Recognition GuideLeafs61100% (5)
- Compabloc Heat Exchangers Cut Downtime at Nynäs RefineryDocument2 pagesCompabloc Heat Exchangers Cut Downtime at Nynäs RefinerylaythNo ratings yet
- Micro Bio Disease ListDocument168 pagesMicro Bio Disease Listspiff spacemanNo ratings yet
- En JV Catalogue 2Document56 pagesEn JV Catalogue 2Yudhi Tri IndrawanNo ratings yet
- 4 WP 2013Document8 pages4 WP 2013TUTORKIMNo ratings yet
- Pharmaceutical Microbiology - B.pharmDocument383 pagesPharmaceutical Microbiology - B.pharmkeyurNo ratings yet
- An Experimental Study of AcidizingDocument11 pagesAn Experimental Study of Acidizinggigi17No ratings yet
- NCERT Class 6 Geography Chapter 1 YouTube Lecture HandoutsDocument5 pagesNCERT Class 6 Geography Chapter 1 YouTube Lecture Handoutskalasri100% (1)
- Project Execution ProcedureDocument4 pagesProject Execution Procedurecyno2167% (3)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (403)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (20)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (5)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- 12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosFrom Everand12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosRating: 4.5 out of 5 stars4.5/5 (207)
- Daniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisFrom EverandDaniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisRating: 3.5 out of 5 stars3.5/5 (130)
- Algorithms to Live By: The Computer Science of Human DecisionsFrom EverandAlgorithms to Live By: The Computer Science of Human DecisionsRating: 4.5 out of 5 stars4.5/5 (722)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsFrom EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsNo ratings yet
- Summary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (11)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningFrom EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningRating: 4 out of 5 stars4/5 (3)