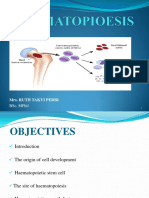
You might also like
- Pathological Processes and Reactive Changes in Leucocyte System Feghiu I., Tacu L.Document13 pagesPathological Processes and Reactive Changes in Leucocyte System Feghiu I., Tacu L.LunguVictoriaNo ratings yet
- Approach To Hematologic DisordersDocument8 pagesApproach To Hematologic DisorderstyasNo ratings yet
- Development and Maintenance of Hematopoietic Tissues: Colony-Forming Units (Cfus)Document13 pagesDevelopment and Maintenance of Hematopoietic Tissues: Colony-Forming Units (Cfus)Tyler KingNo ratings yet
- BLOOD DISEASES: PRODUCTION AND REGULATIONDocument101 pagesBLOOD DISEASES: PRODUCTION AND REGULATIONPaul AshburnerNo ratings yet
- Hematopoiesis and Cells of The Immune SystemDocument19 pagesHematopoiesis and Cells of The Immune SystemCGM BF OFFICENo ratings yet
- Hematopoiesis: Pluri - Several) or Hemocytoblasts. These Cells Have The Capacity To Develop Into Many Different Types ofDocument12 pagesHematopoiesis: Pluri - Several) or Hemocytoblasts. These Cells Have The Capacity To Develop Into Many Different Types ofRajender ArutlaNo ratings yet
- Hemopoiesis PDFDocument14 pagesHemopoiesis PDFrysnawahyu13No ratings yet
- CELLS and ORGANS of IMMUNE SYSTEM - FormattedDocument26 pagesCELLS and ORGANS of IMMUNE SYSTEM - FormattedSantiago MuñozNo ratings yet
- Exercise 5 HematopoiesisDocument11 pagesExercise 5 HematopoiesisAngela AninangNo ratings yet
- BSC Lecture Notes Semester 1Document7 pagesBSC Lecture Notes Semester 1satasha28No ratings yet
- Module Overview: Haematology and Transfusion Science (BMS 303)Document12 pagesModule Overview: Haematology and Transfusion Science (BMS 303)Grace PostmaNo ratings yet
- Lym, Throm, GranuDocument12 pagesLym, Throm, Granumaxwell amponsahNo ratings yet
- Bio 2 Assignment 3Document16 pagesBio 2 Assignment 3Arun RajNo ratings yet
- Lecture 3 (1) HaematopoeisisDocument20 pagesLecture 3 (1) Haematopoeisismaxwell amponsahNo ratings yet
- Semeiology of Hematopoietic SystemDocument18 pagesSemeiology of Hematopoietic SystemIvanNo ratings yet
- Class 12 - FCIA - Composition & Function of Blood - 2022Document5 pagesClass 12 - FCIA - Composition & Function of Blood - 2022Rebeca MedinaNo ratings yet
- TranslateDocument6 pagesTranslatenazarovmuhammad1131No ratings yet
- Leukemia Case StudyDocument10 pagesLeukemia Case StudyQueennieMarelRamos100% (1)
- Bone Marrow and Hematopoiesis - 2Document7 pagesBone Marrow and Hematopoiesis - 2Sharan MurugaboopathyNo ratings yet
- Histologi HematopoeitikDocument29 pagesHistologi Hematopoeitiknia zahroNo ratings yet
- Anatomy and Physiology of LeukemiaDocument7 pagesAnatomy and Physiology of LeukemiaJamaica Aurelio75% (4)
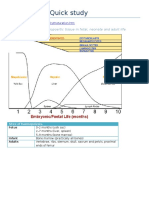
- Hematology Quick StudyDocument47 pagesHematology Quick StudyShanygne Krystal SwannNo ratings yet
- Hemopoiesis1Document17 pagesHemopoiesis1Octafiani Trikartika DarimanNo ratings yet
- Anfisman 2Document2 pagesAnfisman 2Rismalinda LagonahNo ratings yet
- Blood formation: Hematopoiesis and the development of red blood cells, white blood cells, and plateletsDocument24 pagesBlood formation: Hematopoiesis and the development of red blood cells, white blood cells, and plateletshassan aryaniNo ratings yet
- Anatomy and Physiology of Hemopoiesis The Bare Essentials: 1. Stem Cells Must ExistDocument4 pagesAnatomy and Physiology of Hemopoiesis The Bare Essentials: 1. Stem Cells Must ExistThomas Adi KhNo ratings yet
- Hematopoietic Stem CellDocument8 pagesHematopoietic Stem CellMaryam ShahrasbiNo ratings yet
- Nursing Anaphysio ITPDocument8 pagesNursing Anaphysio ITPEköw Santiago JavierNo ratings yet
- Introduction To Haematology and BT 1Document37 pagesIntroduction To Haematology and BT 1akoeljames8543No ratings yet
- Anatomy and physiology of bloodDocument4 pagesAnatomy and physiology of bloodDylle Lorenzo ClaudioNo ratings yet
- PBL Blood DonationDocument5 pagesPBL Blood DonationSamuel GeorgeNo ratings yet
- IUC - Haematology - Lecture 2 - Hemopoiesis-2021Document46 pagesIUC - Haematology - Lecture 2 - Hemopoiesis-2021Kingsly NdangaNo ratings yet
- Case Study - LeukemiaDocument18 pagesCase Study - LeukemiaJerome Valdellon100% (1)
- Lecture 1 - Introduction To HematologyDocument30 pagesLecture 1 - Introduction To Hematologyimam100% (1)
- A Simple Guide to the Blood Cells, Related Diseases And Use in Disease DiagnosisFrom EverandA Simple Guide to the Blood Cells, Related Diseases And Use in Disease DiagnosisNo ratings yet
- Lect 1Document47 pagesLect 1yamanassafNo ratings yet
- Hematology GNDocument94 pagesHematology GNJan Lian Enriquez NoolNo ratings yet
- Hematopoietic DevelopmentDocument6 pagesHematopoietic DevelopmentSea ClestNo ratings yet
- Erythropoiesis: Presented by DR Etu-Efeotor T. PDocument88 pagesErythropoiesis: Presented by DR Etu-Efeotor T. PPrincewill SeiyefaNo ratings yet
- Hematopoiesis, Including Lymphocyte Development and MaturationDocument2 pagesHematopoiesis, Including Lymphocyte Development and MaturationMuhammad FakhrizalNo ratings yet
- CiullahemaDocument60 pagesCiullahemaMariel AbatayoNo ratings yet
- Hematopoiesis New 161214142246Document29 pagesHematopoiesis New 161214142246Vikram KumarNo ratings yet
- Blood SciencesDocument47 pagesBlood Sciencescaroline vaughanNo ratings yet
- Hema TransesDocument25 pagesHema TransesNikoh Anthony EwayanNo ratings yet
- Cell DifferentiationDocument16 pagesCell DifferentiationHarika KeshavNo ratings yet
- Blood CellDocument2 pagesBlood CellJo Louis CamiguingNo ratings yet
- Stem CellDocument8 pagesStem CellHanumat SinghNo ratings yet
- Chapter Iii - Discussion of The DiseaseDocument13 pagesChapter Iii - Discussion of The DiseaseJemima LaigoNo ratings yet
- Seminar Submitted in Partial Fulfillment of Master Degree in Zoology OnDocument24 pagesSeminar Submitted in Partial Fulfillment of Master Degree in Zoology OnKhushbu BhardwajNo ratings yet
- INTRODUCTION TO BLOOD Med st-1Document26 pagesINTRODUCTION TO BLOOD Med st-1Jude ChinecheremNo ratings yet
- FALLSEM2023-24 BBIT305L TH VL2023240100183 2023-04-28 Reference-Material-IDocument9 pagesFALLSEM2023-24 BBIT305L TH VL2023240100183 2023-04-28 Reference-Material-IDrishti BisuiNo ratings yet
- The Anatomy and Functions of BloodDocument14 pagesThe Anatomy and Functions of BloodPari JanNo ratings yet
- 1.anaemia Intro, Retic, Indices-1Document82 pages1.anaemia Intro, Retic, Indices-1InaGargNo ratings yet
- Haemopoiesis: Written By: Fayzah Alshammari Date: 20/02/2021Document5 pagesHaemopoiesis: Written By: Fayzah Alshammari Date: 20/02/2021fayzah alshammariNo ratings yet
- Chapter 20: The Cardiovascular System: Blood: Embryology Atlas HematopoiesisDocument8 pagesChapter 20: The Cardiovascular System: Blood: Embryology Atlas Hematopoiesisfjestefania6892No ratings yet
- Anatomy and Physiology Related To Multiple Myelom1Document15 pagesAnatomy and Physiology Related To Multiple Myelom1Diane Kate Tobias Magno100% (1)
- RBC Lecture Outline Erythropoiesis 2010-01-02Document5 pagesRBC Lecture Outline Erythropoiesis 2010-01-02anil awaseNo ratings yet
- Haematopoitik System: Prof - Dr. Adi Koesoema Aman SPPK (KH) - Dr. Malayana Nasution Mked - SPPKDocument43 pagesHaematopoitik System: Prof - Dr. Adi Koesoema Aman SPPK (KH) - Dr. Malayana Nasution Mked - SPPKrubyniNo ratings yet
- 2016 - Historical and Cultural Aspects of Pineal Gland - Comparison Between The Theories Provided by Spiritism in The 1940s and The Currents Cientific EvidenceDocument12 pages2016 - Historical and Cultural Aspects of Pineal Gland - Comparison Between The Theories Provided by Spiritism in The 1940s and The Currents Cientific Evidencedick_henriqueNo ratings yet
- Specimen 2019 (IAL) MS - Unit 3 Edexcel Biology A-LevelDocument8 pagesSpecimen 2019 (IAL) MS - Unit 3 Edexcel Biology A-Levelchipo activesonNo ratings yet
- Microscopic Organisms Classification QuizDocument8 pagesMicroscopic Organisms Classification QuizAll kinds of information on channelNo ratings yet
- Cambridge IGCSE ™: Biology 0610/41Document12 pagesCambridge IGCSE ™: Biology 0610/41shabanaNo ratings yet
- Forman Christian CollegeDocument2 pagesForman Christian CollegeBhinderNo ratings yet
- Biot Yg I Biotime Fluorescence Immunoassay AnalyzerDocument2 pagesBiot Yg I Biotime Fluorescence Immunoassay Analyzerxuseen maxamedNo ratings yet
- PCR Troubleshooting-Part 1 "No Bands": Hints, Tips and Trouble Shooting For Molecular Biology TechniciansDocument6 pagesPCR Troubleshooting-Part 1 "No Bands": Hints, Tips and Trouble Shooting For Molecular Biology Technicianssam911101No ratings yet
- Dietary Recall Analysis for Underweight PatientDocument7 pagesDietary Recall Analysis for Underweight PatientRayyan HafizNo ratings yet
- BrainAge ConceptsDocument2 pagesBrainAge ConceptsManuela Martín-BejaranoNo ratings yet
- BNWSatire of a Dystopian FutureDocument19 pagesBNWSatire of a Dystopian FutureLudovica MomiNo ratings yet
- Biochemistry TA Notes Part 1Document49 pagesBiochemistry TA Notes Part 1lovelyc95No ratings yet
- The Learners Demonstrate An Understanding Of: The Learners Shall Be Able To: The Learners: The LearnersDocument2 pagesThe Learners Demonstrate An Understanding Of: The Learners Shall Be Able To: The Learners: The LearnersFerdinand GarciaNo ratings yet
- Learner Guide: Cambridge O Level Biology 5090Document45 pagesLearner Guide: Cambridge O Level Biology 5090Dawood100% (1)
- Q1 SHS UCSP Module 4Document23 pagesQ1 SHS UCSP Module 4vience banzaliNo ratings yet
- Csec Biology Manual of Essential Notes Based Purely On Syllabus RequirementsDocument126 pagesCsec Biology Manual of Essential Notes Based Purely On Syllabus RequirementsNicayne RamnarineNo ratings yet
- FINAL Syllabus Changes in Cambridge IGCSE™ Biology (0610) Author LetterDocument2 pagesFINAL Syllabus Changes in Cambridge IGCSE™ Biology (0610) Author LetterAGS GamingNo ratings yet
- Tugas B.inggrisDocument13 pagesTugas B.inggrisDande lionNo ratings yet
- Estuarine, Coastal and Shelf Science: A B A ADocument9 pagesEstuarine, Coastal and Shelf Science: A B A ACarlos SDNo ratings yet
- DARDEN Et Al Reasoning in Biological Discoveries 2006Document372 pagesDARDEN Et Al Reasoning in Biological Discoveries 2006Gustavo CaponiNo ratings yet
- American Survival Guide - June 2015Document132 pagesAmerican Survival Guide - June 2015lucamorlando100% (2)
- EED 480NA STEP TemplateDocument22 pagesEED 480NA STEP TemplateTami GoddardNo ratings yet
- CalCOFI-1981-Rpt Vol 22 1981Document160 pagesCalCOFI-1981-Rpt Vol 22 1981A. Miguel SantosNo ratings yet
- Analisis Molekuler Tingkat Kekerabatan Babi Hutan Dan Babi RusaDocument12 pagesAnalisis Molekuler Tingkat Kekerabatan Babi Hutan Dan Babi Rusanaura ANo ratings yet
- Chapter 1.0: Biodiversity: of Different Things. A Given AreaDocument61 pagesChapter 1.0: Biodiversity: of Different Things. A Given AreaNURUL HIDAYAH BINTI SAIFUL ANUAR MoeNo ratings yet
- Dr. Sudha Kumari Assistant Professor Department of Veterinary Microbiology Bihar Animal Sciences University, PatnaDocument15 pagesDr. Sudha Kumari Assistant Professor Department of Veterinary Microbiology Bihar Animal Sciences University, PatnaShafici CqadirNo ratings yet
- Citrus Commodity Survey ReferenceDocument255 pagesCitrus Commodity Survey Referenceduong-hoai-an-6140No ratings yet
- Matta Et Al. - 2018 - Development and Characterization of A Novel LucifeDocument14 pagesMatta Et Al. - 2018 - Development and Characterization of A Novel LucifeRaghav VijNo ratings yet
- The Selection Process WorksheetDocument2 pagesThe Selection Process Worksheetapi-256236481No ratings yet
- Virtual Lab Report: Name of The SimulationDocument12 pagesVirtual Lab Report: Name of The SimulationBruce PanNo ratings yet
- SOWDAMBIKAA GROUP OF SCHOOLS BIOLOGY SLIP TESTDocument2 pagesSOWDAMBIKAA GROUP OF SCHOOLS BIOLOGY SLIP TESTfireNo ratings yet