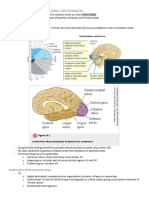
You might also like
- Guide To The Dissection of The Dog, 8th EditionDocument341 pagesGuide To The Dissection of The Dog, 8th EditionMaisy Rommela MedallaNo ratings yet
- Exercise 4 Frog Embryo 4mm 7mm 10mmDocument23 pagesExercise 4 Frog Embryo 4mm 7mm 10mmrexartooz95% (19)
- Zeki Disunity of ConsciousnessDocument5 pagesZeki Disunity of ConsciousnessRose DarugarNo ratings yet
- Visual Cortex - WikipediaDocument23 pagesVisual Cortex - WikipediaMM NabeelNo ratings yet
- Reticular FormationDocument26 pagesReticular Formationsr_sushilNo ratings yet
- Armaturi Profile PVCDocument53 pagesArmaturi Profile PVCstoian andreiNo ratings yet
- Exercise Induced Neuroplasticity - Rogge 2018Document38 pagesExercise Induced Neuroplasticity - Rogge 2018Juani Cantellanos0% (1)
- The Asynchrony of ConsciousnessDocument3 pagesThe Asynchrony of ConsciousnessstefgerosaNo ratings yet
- Parallel Processing, Asynchronous Perception, and A Distributed System of Consciousness in VisionDocument8 pagesParallel Processing, Asynchronous Perception, and A Distributed System of Consciousness in VisionRodrigo GoulartNo ratings yet
- Distributed Hierarchical Processing in The Primate Cerebral CortexDocument47 pagesDistributed Hierarchical Processing in The Primate Cerebral CortexfrereretyNo ratings yet
- Primary Visual Cortex Activity Along The Apparent-Motion Trace Reflects Illusory PerceptionDocument10 pagesPrimary Visual Cortex Activity Along The Apparent-Motion Trace Reflects Illusory PerceptionFrontiersNo ratings yet
- Jesse J. Prinz (2000), A Neurofunctional Theory of Visual ConsciousnessDocument17 pagesJesse J. Prinz (2000), A Neurofunctional Theory of Visual ConsciousnessRogério P. SeveroNo ratings yet
- The Next Steps in Visual Processing: An Experiment On Colour AdaptationDocument1 pageThe Next Steps in Visual Processing: An Experiment On Colour AdaptationVipin ValsalanNo ratings yet
- My DLunit 2 FRM TXTDocument18 pagesMy DLunit 2 FRM TXTVali BhashaNo ratings yet
- TMP 5 FADocument12 pagesTMP 5 FAFrontiersNo ratings yet
- Psych 1XX3 Form Perception II Lecture NotesDocument11 pagesPsych 1XX3 Form Perception II Lecture NotesGurry MannNo ratings yet
- Tmp18a6 TMPDocument25 pagesTmp18a6 TMPFrontiersNo ratings yet
- Two Neural Correlates of ConsciousnessDocument7 pagesTwo Neural Correlates of ConsciousnessCuirocamposNo ratings yet
- (2011) The Organization and DissolutionDocument11 pages(2011) The Organization and Dissolutionniles13No ratings yet
- Two Neural Correlates of ConsciousnessDocument7 pagesTwo Neural Correlates of ConsciousnessKapderGutenHoffnungNo ratings yet
- International Phenomenological SocietyDocument7 pagesInternational Phenomenological SocietyCamilo Torres MestraNo ratings yet
- MirrorBot Report Localises Objects Using Neural Associator NetworkDocument20 pagesMirrorBot Report Localises Objects Using Neural Associator NetworkMahesh Kumar GubbalaNo ratings yet
- Conscious, Preconscious, and Subliminal Processing: A Testable TaxonomyDocument8 pagesConscious, Preconscious, and Subliminal Processing: A Testable TaxonomyMica Ballito de TroyaNo ratings yet
- Do We Know What The Early Visual System Does?: Mini-SymposiumDocument21 pagesDo We Know What The Early Visual System Does?: Mini-SymposiumShah JamalNo ratings yet
- Prinz Neuro Functional To CDocument11 pagesPrinz Neuro Functional To CHipatia AlNo ratings yet
- Ungerleider & Bell (2011) - Uncovering The Visual Alphabet. Advances in Our Understanding PDFDocument18 pagesUngerleider & Bell (2011) - Uncovering The Visual Alphabet. Advances in Our Understanding PDFJhamil Andre Venero LovatonNo ratings yet
- CH 3Document8 pagesCH 3Sabrina MaciarielloNo ratings yet
- C81BIO - Occipital and Temporal LobeDocument24 pagesC81BIO - Occipital and Temporal LobeMelissa 'meli' LapianNo ratings yet
- tmpCD34 TMPDocument13 pagestmpCD34 TMPFrontiersNo ratings yet
- Separate Visual Pathways For Perception and Action (Goodale, Milner)Document6 pagesSeparate Visual Pathways For Perception and Action (Goodale, Milner)C.No ratings yet
- Visual Perception: The Merging of the Visible and IntellectualDocument11 pagesVisual Perception: The Merging of the Visible and IntellectualJanina VienišėNo ratings yet
- Conscious Visual PerceptionDocument46 pagesConscious Visual PerceptionDa CuNo ratings yet
- 1 s2.0 S1053811921008971 MainDocument17 pages1 s2.0 S1053811921008971 Mainspamkt2002No ratings yet
- Chapter 13 The Occipital Lobes and NetworksDocument8 pagesChapter 13 The Occipital Lobes and NetworkstaniaNo ratings yet
- 1 s2.0 S0167876015001865 MainDocument11 pages1 s2.0 S0167876015001865 Mainsantosh kumarNo ratings yet
- VisualDocument3 pagesVisualMelisa HilaireNo ratings yet
- Color-Capable Neocognitron: Mechanical Engineering Department, American University of Beirut, Beirut, LebanonDocument30 pagesColor-Capable Neocognitron: Mechanical Engineering Department, American University of Beirut, Beirut, Lebanondanny asmarNo ratings yet
- Apontamentos de Perceção e AtençãoDocument3 pagesApontamentos de Perceção e AtençãoMarianaNo ratings yet
- Clark - Unravelling The Golden BraidDocument9 pagesClark - Unravelling The Golden BraidDavid_Silverma_8933No ratings yet
- Rolfs Perception 2015Document20 pagesRolfs Perception 2015lorelenNo ratings yet
- Biol Cybern (2015) 109:215-239 STDP in Lateral Connections Creates Category-Based Perceptual Cycles For Invariance Learning With Multiple StimuliDocument25 pagesBiol Cybern (2015) 109:215-239 STDP in Lateral Connections Creates Category-Based Perceptual Cycles For Invariance Learning With Multiple StimuliOleg NikitinNo ratings yet
- Visual Processing System in the BrainDocument15 pagesVisual Processing System in the BrainAbhishek WarokarNo ratings yet
- Retina-inspired filter extracts visual infoDocument13 pagesRetina-inspired filter extracts visual infoAkash SaxenaNo ratings yet
- Deep Learning CNNs ExplainedDocument32 pagesDeep Learning CNNs ExplainedQ QNo ratings yet
- Convolutional Neural Networks: Fundamentals and Applications for Analyzing Visual ImageryFrom EverandConvolutional Neural Networks: Fundamentals and Applications for Analyzing Visual ImageryNo ratings yet
- György Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)Document20 pagesGyörgy Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)juannnnNo ratings yet
- An Introduction To Deep Learning: January 2011Document14 pagesAn Introduction To Deep Learning: January 2011TekNo ratings yet
- Disentangling Conscious From Unconscious Cognitive Processing With Event-Related EEG PotentialsDocument8 pagesDisentangling Conscious From Unconscious Cognitive Processing With Event-Related EEG PotentialsTheGeeks LegendNo ratings yet
- Visual Mental Imagery: What The Head's Eye Tells The Mind's EyeDocument11 pagesVisual Mental Imagery: What The Head's Eye Tells The Mind's Eyeeli_wenNo ratings yet
- BMB 413 Neuroscience Online Notes Vol4Document19 pagesBMB 413 Neuroscience Online Notes Vol4Moose KeNo ratings yet
- Visual Perception of Spatial SubjectsDocument8 pagesVisual Perception of Spatial SubjectsvrapciudorianNo ratings yet
- How The Brain WorksDocument7 pagesHow The Brain WorksWy TeayNo ratings yet
- Gabriel Axel - Master Thesis - Vision The Art of Evolution (2010)Document41 pagesGabriel Axel - Master Thesis - Vision The Art of Evolution (2010)gabrielaxelNo ratings yet
- Carl R Olson: Object-Based Vision and Attention in PrimatesDocument9 pagesCarl R Olson: Object-Based Vision and Attention in PrimatesPablo Ignacio Nova BaezaNo ratings yet
- Consciousness in The Universe Is Scale Invariant and Implies An Event Horizon of The Human Brain PDFDocument39 pagesConsciousness in The Universe Is Scale Invariant and Implies An Event Horizon of The Human Brain PDFAdrián GonzálezNo ratings yet
- Imaging Brain Function With EEG Advanced Temporal and Spatial Analysis of Electroencephalographic Signals 2012Document267 pagesImaging Brain Function With EEG Advanced Temporal and Spatial Analysis of Electroencephalographic Signals 2012Rebeca Pardo GarcíaNo ratings yet
- Vergence Control in A Binocular Vision System Using Weightless Neural NetworksDocument8 pagesVergence Control in A Binocular Vision System Using Weightless Neural NetworksprofakarinNo ratings yet
- Gollisch MeisterDocument15 pagesGollisch MeisterElaheh LotfiNo ratings yet
- György Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)Document20 pagesGyörgy Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)juannnnNo ratings yet
- 1992,Distal attribution and presence(引用518)Document7 pages1992,Distal attribution and presence(引用518)bingwan zhangNo ratings yet
- Review: Conscious Processing and The Global Neuronal Workspace HypothesisDocument23 pagesReview: Conscious Processing and The Global Neuronal Workspace HypothesisSebastian Gonzalez MazoNo ratings yet
- 2005 - Baars - Global Workspace Theory of Consciousness - Toward A Cognitive Neuroscience of Human ExperienceDocument9 pages2005 - Baars - Global Workspace Theory of Consciousness - Toward A Cognitive Neuroscience of Human ExperiencefachavalNo ratings yet
- Neurophysiology of Binocular VisionDocument7 pagesNeurophysiology of Binocular Visionitay778No ratings yet
- Chapter 2 NS Objective AnswersDocument6 pagesChapter 2 NS Objective AnswersSadasivarao GaddeNo ratings yet
- Chapter-5.-Vision (3)Document33 pagesChapter-5.-Vision (3)yamquim01No ratings yet
- GooglepreviewDocument12 pagesGooglepreviewCelia VillodreNo ratings yet
- 5 Ardila Historia Agnosias Enero Junio Vol 151 2015Document8 pages5 Ardila Historia Agnosias Enero Junio Vol 151 2015Celia Villodre LasernaNo ratings yet
- Agnosias: Wiley Interdisciplinary Reviews. Cognitive Science March 2010Document12 pagesAgnosias: Wiley Interdisciplinary Reviews. Cognitive Science March 2010Celia VillodreNo ratings yet
- Agnosias: Wiley Interdisciplinary Reviews. Cognitive Science March 2010Document12 pagesAgnosias: Wiley Interdisciplinary Reviews. Cognitive Science March 2010Celia VillodreNo ratings yet
- Friederici - 2013 - The Language Network PDFDocument5 pagesFriederici - 2013 - The Language Network PDFHansel SotoNo ratings yet
- Anatomie GrileDocument96 pagesAnatomie GrileMarin TheodorNo ratings yet
- Dopamine Pathways PDFDocument3 pagesDopamine Pathways PDFMuhammad Zul Fahmi AkbarNo ratings yet
- Congenital Malformation of Nervous System PPT Sesi 1Document22 pagesCongenital Malformation of Nervous System PPT Sesi 1Brandedlovers Onlineshop100% (1)
- Battery CodesDocument3 pagesBattery CodesWalter MaqueraNo ratings yet
- Birth Asphyxia: DefinitionDocument5 pagesBirth Asphyxia: DefinitionDella FerginaNo ratings yet
- The Brain and Learning WorksheetDocument5 pagesThe Brain and Learning WorksheetJayRAjNo ratings yet
- Learning Theory of NeuroscienceDocument8 pagesLearning Theory of Neuroscienceazahra hardi cusiniaNo ratings yet
- What Is The Brain?: CerebrumDocument3 pagesWhat Is The Brain?: CerebrumAurea Jasmine DacuycuyNo ratings yet
- Language and The BrainDocument8 pagesLanguage and The BrainAntonio GoqueNo ratings yet
- S05.s2 - Lectura-Estructura Anatomofuncional Del NeocortexDocument23 pagesS05.s2 - Lectura-Estructura Anatomofuncional Del Neocortexhpamela91No ratings yet
- 1.0 Basic Brain Anatomy CDocument9 pages1.0 Basic Brain Anatomy Cryanpelletier0No ratings yet
- NBSS CASE 6 - Cerebral Palsy and Mental RetardationDocument28 pagesNBSS CASE 6 - Cerebral Palsy and Mental RetardationbungadielaNo ratings yet
- Cleavage Formation and Blastulation: Lesson 2.3: Animal DevelopmentDocument8 pagesCleavage Formation and Blastulation: Lesson 2.3: Animal DevelopmentWestminster AbbeyNo ratings yet
- Ibtl Lec 16 PDFDocument7 pagesIbtl Lec 16 PDFgreen1458No ratings yet
- Anatomy and Physiology of the BrainDocument13 pagesAnatomy and Physiology of the BrainEnna PaulaNo ratings yet
- Lecture 9 - Intro To Memory & Encoding - FullDocument18 pagesLecture 9 - Intro To Memory & Encoding - FullJoshNo ratings yet
- Brain Anatomy - Dr. Richard Yan Marvellini, SpRadDocument32 pagesBrain Anatomy - Dr. Richard Yan Marvellini, SpRadTahu Tempe100% (1)
- What Does It Do?: What Happens When The Temporal Lobe Is Damaged?Document2 pagesWhat Does It Do?: What Happens When The Temporal Lobe Is Damaged?Tu Thanh LuongNo ratings yet
- Human Memory and The Medial Temporal Region of The Brain PDFDocument84 pagesHuman Memory and The Medial Temporal Region of The Brain PDFIcaroNo ratings yet
- Module 5 & 6Document36 pagesModule 5 & 6joyNo ratings yet
- Frog Embryo Germ Layer DevelopmentDocument10 pagesFrog Embryo Germ Layer DevelopmentJervis PendletonNo ratings yet
- Cortical and Subcortical Areas of The BrainDocument1 pageCortical and Subcortical Areas of The BrainMahesh DayalNo ratings yet
- Superior Longitudinal Fasciculus - Connectopedia Knowledge Database - PR Denis DucreuxDocument3 pagesSuperior Longitudinal Fasciculus - Connectopedia Knowledge Database - PR Denis DucreuxpagelerNo ratings yet
- 075 - Neurology Physiology) Cerebrum - Temporal Lobe Anatomy & FunctionDocument2 pages075 - Neurology Physiology) Cerebrum - Temporal Lobe Anatomy & Functionzayat13No ratings yet