You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
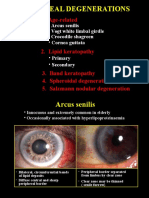
- 16corneal DegenerationsDocument11 pages16corneal DegenerationsmaulanaNo ratings yet
- MasterSAT 1056 Pages 2-1056 592-594, 668-670, 761-763Document9 pagesMasterSAT 1056 Pages 2-1056 592-594, 668-670, 761-763Kumer Shinfa0% (1)
- TherapyDocument25 pagesTherapyHagai MagaiNo ratings yet
- Anchorage in OrthodonticsDocument25 pagesAnchorage in OrthodonticsJavariya AshrafNo ratings yet
- AdmittanceCard PDFDocument1 pageAdmittanceCard PDFJavariya AshrafNo ratings yet
- Class I MalocclusionDocument24 pagesClass I MalocclusionJavariya AshrafNo ratings yet
- Bds Third Professional Examination 2007 General Medicine Table of Specifications Chapter Wise Distribution of Mcqs and SeqsDocument5 pagesBds Third Professional Examination 2007 General Medicine Table of Specifications Chapter Wise Distribution of Mcqs and SeqsJavariya AshrafNo ratings yet
- Sexually Transmitted Diseases OverviewDocument45 pagesSexually Transmitted Diseases OverviewJean De Vera MelendezNo ratings yet
- Physical GrowthDocument5 pagesPhysical GrowthrevgandaNo ratings yet
- Drugs Affecting The Central Nervous SystemDocument22 pagesDrugs Affecting The Central Nervous Systemአብይ በላይነሽ ጥላሁንNo ratings yet
- Full Download Book Ryans Retina PDFDocument41 pagesFull Download Book Ryans Retina PDFdavid.gayer737100% (20)
- Handbook of Emergency MedicineDocument298 pagesHandbook of Emergency MedicineJazlan Mohamad100% (15)
- Assessment MnemonicsDocument23 pagesAssessment MnemonicsAilene Ponce Fillon100% (1)
- Non Sirosis PHDocument11 pagesNon Sirosis PHHIstoryNo ratings yet
- Color Therapy TreatmentDocument30 pagesColor Therapy TreatmentDaniel ChrysNo ratings yet
- The Impact of Overconsumption of SugarDocument11 pagesThe Impact of Overconsumption of SugarHermilinda Orenze100% (5)
- Nerve Suply To Head and NeckDocument22 pagesNerve Suply To Head and NeckrajtanniruNo ratings yet
- Valvular Heart Disease To TW FinalDocument13 pagesValvular Heart Disease To TW FinalMohammed ElSayedNo ratings yet
- Angina PectorisDocument17 pagesAngina PectorisRakesh Reddy100% (1)
- Management of CataractDocument25 pagesManagement of Cataractapi-3742497No ratings yet
- Cdmp2015 HandbookDocument90 pagesCdmp2015 HandbookTony NgNo ratings yet
- Professional Development Resource - Promoting Best Practice: "Palliative Care Under Observaton."Document8 pagesProfessional Development Resource - Promoting Best Practice: "Palliative Care Under Observaton."karma SherpaNo ratings yet
- The Standard 30.04.2014Document80 pagesThe Standard 30.04.2014Michelle WhiteNo ratings yet
- Pre-Op Assessment: Dr. Ayesha Ehsan House Surgeon S-IIDocument25 pagesPre-Op Assessment: Dr. Ayesha Ehsan House Surgeon S-IIFatima MNo ratings yet
- Field Trip Permission FormDocument2 pagesField Trip Permission FormRubylyn Dela PenaNo ratings yet
- NCM 104 Indiv and FamilyDocument123 pagesNCM 104 Indiv and FamilyJames Peter ManatadNo ratings yet
- Brain DamageDocument5 pagesBrain DamageAzra KurtovićNo ratings yet
- Assessing Solutions/To Prevent Depression (2019) : Pavia National High School Evangelista ST., Pavia, IloiloDocument13 pagesAssessing Solutions/To Prevent Depression (2019) : Pavia National High School Evangelista ST., Pavia, IloiloChimmy NochuNo ratings yet
- Homocysteine and Markers of Coagulation and Endothelial Cell ActivationDocument7 pagesHomocysteine and Markers of Coagulation and Endothelial Cell ActivationMaya RustamNo ratings yet
- Endodontic: EmergenciesDocument53 pagesEndodontic: EmergenciesAhmed S. Abu-Haimed100% (1)
- Scolioza CongenitalaDocument20 pagesScolioza CongenitalaIulia Dulgheru100% (1)
- Rearrange The Jumble Letters: 10 1 10Document4 pagesRearrange The Jumble Letters: 10 1 10Shafee ArafatNo ratings yet
- Surgical Notes (Teddy)Document88 pagesSurgical Notes (Teddy)shichianNo ratings yet
- Bethlehem University Faculty of Nursing and Health Science Psychiatric Mental Health Nursing NURS 342Document24 pagesBethlehem University Faculty of Nursing and Health Science Psychiatric Mental Health Nursing NURS 342Yousef JafarNo ratings yet