You might also like
- Chapman 1950Document5 pagesChapman 1950poojajain86No ratings yet
- Prevention of Shedding and Re-Shedding of Toxoplasma Gondii in Cats. Malmasi, 2008Document3 pagesPrevention of Shedding and Re-Shedding of Toxoplasma Gondii in Cats. Malmasi, 2008Ivan MQNo ratings yet
- Influence of Subtherapeutic Levels of Oxytetacycline On Salmonella Typhimurim in Swine, Calves, and ChickensDocument9 pagesInfluence of Subtherapeutic Levels of Oxytetacycline On Salmonella Typhimurim in Swine, Calves, and ChickensLa Fille D'ÈveNo ratings yet
- Journal ResearchDocument4 pagesJournal ResearchFaizaSenjaWidyaPerdanaNo ratings yet
- FleascontrolDocument8 pagesFleascontrolVT BabuNo ratings yet
- Tuberculostatic Activity of Henna (LawsoniaDocument3 pagesTuberculostatic Activity of Henna (Lawsonia9884288233No ratings yet
- Imunomodulatory PhytotherapyDocument7 pagesImunomodulatory PhytotherapyMuddasser JamalNo ratings yet
- The American Society of ParasitologistsDocument5 pagesThe American Society of ParasitologistsangelNo ratings yet
- Daugschies 2000Document3 pagesDaugschies 2000Ivan MQNo ratings yet
- Research Article in Vivo Antimalarial Activity of Annona Muricata Leaf Extract Plasmodium BergheiDocument5 pagesResearch Article in Vivo Antimalarial Activity of Annona Muricata Leaf Extract Plasmodium BergheifikirudinNo ratings yet
- Worm and Larval Burden, Histopathological and Ultrastructural Evaluation of T. Spiralis Vaccination Using Crude Worms And/or Larvae Antigens: Experimental StudiesDocument12 pagesWorm and Larval Burden, Histopathological and Ultrastructural Evaluation of T. Spiralis Vaccination Using Crude Worms And/or Larvae Antigens: Experimental StudiesSari RamadhanNo ratings yet
- Duration of Protective Immunity Against Ovine Haemonchosis Following Vaccination With The Nematode Gut Membrane Antigen H L LDocument5 pagesDuration of Protective Immunity Against Ovine Haemonchosis Following Vaccination With The Nematode Gut Membrane Antigen H L LAngéllica MaríaNo ratings yet
- From The Istituto Nazionale Per Lo Studio e La Cura Dei Tumori, 20133milano, ItalyDocument9 pagesFrom The Istituto Nazionale Per Lo Studio e La Cura Dei Tumori, 20133milano, Italyniken retnoNo ratings yet
- Pasteurellamultocida Type DDocument4 pagesPasteurellamultocida Type Dali hendyNo ratings yet
- 159 FullDocument19 pages159 FullMina BekNo ratings yet
- Potensi Anthelmintik Akar Tanaman Putri Malu (Mimosa Pudica L.) Terhadap Hymenolepis Nana Pada MencitDocument7 pagesPotensi Anthelmintik Akar Tanaman Putri Malu (Mimosa Pudica L.) Terhadap Hymenolepis Nana Pada MencitJulfaind HammaNo ratings yet
- Yardley y Croft, 1999Document5 pagesYardley y Croft, 1999jesusNo ratings yet
- 1 s2.0 S222116911530109X MainDocument5 pages1 s2.0 S222116911530109X MainSantosh GejageNo ratings yet
- Study of The Role of Pigeons in The Dissemination of Cryptococcus Neoformans in NatureDocument6 pagesStudy of The Role of Pigeons in The Dissemination of Cryptococcus Neoformans in NatureTANIA CORONADO ROMERONo ratings yet
- Prophylactic Efficacy of Some Antibiotic Combinations Against Brucella MelitensisDocument10 pagesProphylactic Efficacy of Some Antibiotic Combinations Against Brucella MelitensisKraken UrNo ratings yet
- 10 1136@vr 147 25 709Document5 pages10 1136@vr 147 25 709Erick ManosalvasNo ratings yet
- Anthelmintic Activity of Praziquantel On PDFDocument2 pagesAnthelmintic Activity of Praziquantel On PDFFinishNo ratings yet
- Preliminary Report On The Clinical and HematologicalDocument28 pagesPreliminary Report On The Clinical and Hematological99jjNo ratings yet
- Parasitic Nematodes MexicoDocument5 pagesParasitic Nematodes MexicoTlaloc ConsultantsNo ratings yet
- Prevalence of Diminazene Aceturate Resistant Trypanosomiasis in Wad Sheep in Plateau State, NigeriaDocument11 pagesPrevalence of Diminazene Aceturate Resistant Trypanosomiasis in Wad Sheep in Plateau State, NigeriaInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Abdel Salam2014Document6 pagesAbdel Salam2014LUIGI ORLANDO FREIRE MARTINEZNo ratings yet
- Trichinellosis: C H A P T E R 2 - 1 - 1 6Document9 pagesTrichinellosis: C H A P T E R 2 - 1 - 1 6syafahalimNo ratings yet
- Alleviation Hysteria Laying Dietary Tryptophan: of in Hens WithDocument5 pagesAlleviation Hysteria Laying Dietary Tryptophan: of in Hens WithImelda MertasariNo ratings yet
- Adulticidal and Larvicidal Effects of ImidaclopridDocument3 pagesAdulticidal and Larvicidal Effects of Imidaclopridtaner_soysurenNo ratings yet
- Baba T.1984. Cell-Mediated Immune Protection in Chickens Against P MultocidaDocument6 pagesBaba T.1984. Cell-Mediated Immune Protection in Chickens Against P MultocidakrodriguezNo ratings yet
- 086 SabinaDocument10 pages086 SabinaMahdaNo ratings yet
- Comparative Methods For Demonstration: Study of Tissue Culture of Toxoplasma GondiiDocument4 pagesComparative Methods For Demonstration: Study of Tissue Culture of Toxoplasma GondiiSalim JufriNo ratings yet
- Melatonin 2Document6 pagesMelatonin 2Andreas TassopoulosNo ratings yet
- Antitumor Activity of An Ethanol Extract of Nigella Sativa SeedsDocument6 pagesAntitumor Activity of An Ethanol Extract of Nigella Sativa SeedsRaden WisnugrohoNo ratings yet
- Trichinellosis: C H A P T E R 2 - 1 - 1 6Document9 pagesTrichinellosis: C H A P T E R 2 - 1 - 1 6Andre BroesNo ratings yet
- Vol11 Iss1 BeugnetDocument6 pagesVol11 Iss1 BeugnetSheikh Abdul WahabNo ratings yet
- Mikro PDFDocument6 pagesMikro PDFYudha Pradhana PutraNo ratings yet
- JM pharmacon,+8.+Sri+Gentari+Benjamin (55-62)Document8 pagesJM pharmacon,+8.+Sri+Gentari+Benjamin (55-62)gupro thaiNo ratings yet
- Neal, 1984 PDFDocument5 pagesNeal, 1984 PDFGutierrez Guarnizo SneiderNo ratings yet
- Oral and Intraperitoneal LD of Thymoquinone, An Active Principle of Nigella Sativa, in Mice and RatsDocument3 pagesOral and Intraperitoneal LD of Thymoquinone, An Active Principle of Nigella Sativa, in Mice and Ratsige zaharaNo ratings yet
- BH Chapter 5 Mice-IMI-Model (1) .EditedDocument16 pagesBH Chapter 5 Mice-IMI-Model (1) .EditedGifted MaggieNo ratings yet
- Aggarwal 2011Document8 pagesAggarwal 2011rani nareza ulfaNo ratings yet
- Antibacterial Activity and in Vitro Anti-Tumor Activity of The Extract of The Larvae of The Housefly (Musca Domestica)Document5 pagesAntibacterial Activity and in Vitro Anti-Tumor Activity of The Extract of The Larvae of The Housefly (Musca Domestica)metabolismeproteinNo ratings yet
- Tinospora Cordifolia: Enhanced Phagocytosis and Antibody Production by - A New Dimension in ImmunomodulationDocument5 pagesTinospora Cordifolia: Enhanced Phagocytosis and Antibody Production by - A New Dimension in ImmunomodulationAlna TechnicalNo ratings yet
- Veterinary Immunology and ImmunopathologyDocument8 pagesVeterinary Immunology and ImmunopathologyMatthew HsuNo ratings yet
- ChikDocument6 pagesChikNtra WahyuNo ratings yet
- Efficacy of Treatments With Toltrazuril 7.5% and Lasalocid Sodium in Sheep Naturally Infected With Eimeria SPPDocument6 pagesEfficacy of Treatments With Toltrazuril 7.5% and Lasalocid Sodium in Sheep Naturally Infected With Eimeria SPPAli ValenciaNo ratings yet
- Islet Cell Aislamiento en Experimental D, L-Etionina Pancreatitis en Perro1-S2.0-0022480485900381-MainDocument9 pagesIslet Cell Aislamiento en Experimental D, L-Etionina Pancreatitis en Perro1-S2.0-0022480485900381-MainCarlos VargasNo ratings yet
- 093 Salmonellosis PDFDocument3 pages093 Salmonellosis PDFMariana MansillaNo ratings yet
- Deken Et Al, 1994 (MAb)Document5 pagesDeken Et Al, 1994 (MAb)Farkhanda SadafNo ratings yet
- Flavonoids From Boldoa Purpurascens Inhibit ProinflammatoryDocument5 pagesFlavonoids From Boldoa Purpurascens Inhibit ProinflammatoryJOHN HELIBERTH ANDREY TORRES GRANADOSNo ratings yet
- Common Effects of Mefenamic AcidDocument4 pagesCommon Effects of Mefenamic AcidAries Templo TaliquigNo ratings yet
- Anesthesiologists 26th AnnualMeeting New Orleans October 2001Document16 pagesAnesthesiologists 26th AnnualMeeting New Orleans October 2001Reginaldo CunhaNo ratings yet
- Avila Et Al-2016-Parasite ImmunologyDocument5 pagesAvila Et Al-2016-Parasite ImmunologyVictória PanassoloNo ratings yet
- The Effects of Photoperiod Programs On Broiler Chicken Performance and Immune ResponseDocument7 pagesThe Effects of Photoperiod Programs On Broiler Chicken Performance and Immune ResponseFlor SagnipNo ratings yet
- IMM 265 - Galley Proof - 5may21 - VelikkakamDocument13 pagesIMM 265 - Galley Proof - 5may21 - VelikkakamSoraya Torres GazeNo ratings yet
- Trichinellosis: C H A P T E R 2 - 1 - 1 6Document9 pagesTrichinellosis: C H A P T E R 2 - 1 - 1 6WormInchNo ratings yet
- Toltrazuril (Baycox) Treatment Against Coccidiosis Caused by Eimeria Sp. in Japanese Quails (Coturnix Coturnix Japonica)Document4 pagesToltrazuril (Baycox) Treatment Against Coccidiosis Caused by Eimeria Sp. in Japanese Quails (Coturnix Coturnix Japonica)khaled piksouNo ratings yet
- The Toxicity of Caffein: An experimental study on different species of animalsFrom EverandThe Toxicity of Caffein: An experimental study on different species of animalsNo ratings yet
- Music Example 7Document6 pagesMusic Example 7Rezita RamadhaniNo ratings yet
- Music Answers 6Document6 pagesMusic Answers 6Rezita RamadhaniNo ratings yet
- Music Demo 2Document5 pagesMusic Demo 2Rezita RamadhaniNo ratings yet
- Music Diagram 1Document7 pagesMusic Diagram 1Rezita RamadhaniNo ratings yet
- Music Demo 8Document6 pagesMusic Demo 8Rezita RamadhaniNo ratings yet
- Music Cheat Sheet 10Document5 pagesMusic Cheat Sheet 10Rezita RamadhaniNo ratings yet
- Music Answers 4Document5 pagesMusic Answers 4Rezita RamadhaniNo ratings yet
- Contents CLDDocument4 pagesContents CLDRezita RamadhaniNo ratings yet
- Music Exercise 6Document5 pagesMusic Exercise 6Rezita RamadhaniNo ratings yet
- Music Lesson Plan 1Document6 pagesMusic Lesson Plan 1Rezita RamadhaniNo ratings yet
- Preface CLDDocument2 pagesPreface CLDRezita RamadhaniNo ratings yet
- Contributors CLDDocument3 pagesContributors CLDRezita RamadhaniNo ratings yet
- Index CLDDocument14 pagesIndex CLDRezita RamadhaniNo ratings yet
- Eleanor FitzgeraldDocument1 pageEleanor FitzgeraldRezita RamadhaniNo ratings yet
- 1939 4551 1 9 145Document11 pages1939 4551 1 9 145Rezita RamadhaniNo ratings yet
- 1 SM PDFDocument12 pages1 SM PDFYoga Maulana RahmanNo ratings yet
- EFEK TONIK EKSTRAK DAUN KENIKIR (Cosmos Caudatus Kunth.) Serta Sumbangannya Pada Pembelajaran Biologi SmaDocument10 pagesEFEK TONIK EKSTRAK DAUN KENIKIR (Cosmos Caudatus Kunth.) Serta Sumbangannya Pada Pembelajaran Biologi SmaMudhakiyah ZaqiatulNo ratings yet
- A RTIKELDocument20 pagesA RTIKELrizky fahrezaNo ratings yet
- 1939 4551 1 9 145Document11 pages1939 4551 1 9 145Rezita RamadhaniNo ratings yet
- Journal of Industrial and Engineering Chemistry: Silvia Curteanu, Mirela Smarandoiu, Doina Horoba, Florin LeonDocument4 pagesJournal of Industrial and Engineering Chemistry: Silvia Curteanu, Mirela Smarandoiu, Doina Horoba, Florin LeonRezita RamadhaniNo ratings yet
- The Concept of Sublimation - Iodine As An Example: Marina Stojanovska, Vladimir M. Petruševski, Bojan ŠoptrajanovDocument5 pagesThe Concept of Sublimation - Iodine As An Example: Marina Stojanovska, Vladimir M. Petruševski, Bojan ŠoptrajanovRezita RamadhaniNo ratings yet
- 05.70.0072 Ivonne Kristina Hidajat Sulistio - DAFTAR PUSTAKADocument4 pages05.70.0072 Ivonne Kristina Hidajat Sulistio - DAFTAR PUSTAKARezita RamadhaniNo ratings yet
- Silver MSDSDocument21 pagesSilver MSDSTueNo ratings yet
- Hortatory ExpositionDocument1 pageHortatory ExpositionRezita RamadhaniNo ratings yet
- Cancer DevelopmentDocument9 pagesCancer DevelopmentRezita RamadhaniNo ratings yet
- 1Document1 page1Rezita RamadhaniNo ratings yet
- BeritaDocument2 pagesBeritaRezita RamadhaniNo ratings yet
- Silver MSDSDocument21 pagesSilver MSDSTueNo ratings yet
- Hortatory ExpositionDocument1 pageHortatory ExpositionRezita RamadhaniNo ratings yet
- 91 Clinical ScenariosDocument198 pages91 Clinical Scenarioszph7100% (1)
- Nigella Sativa and Cimetidine Against AcetylsalicylicDocument6 pagesNigella Sativa and Cimetidine Against AcetylsalicylicTeguh D. WicaksonoNo ratings yet
- Basic of AccupressureDocument48 pagesBasic of AccupressureRavikumarrd Ravi100% (2)
- 91598662Document53 pages91598662Ibrahim SiddigNo ratings yet
- Discurssive Writing TopicDocument8 pagesDiscurssive Writing TopicAscensión Villalba VaronaNo ratings yet
- Gynae 64Document14 pagesGynae 64Jha K SujitNo ratings yet
- Nematode StructureDocument31 pagesNematode StructureShadab HanafiNo ratings yet
- 03etiology and Pathogenesis of Neonatal EncephalopathyDocument22 pages03etiology and Pathogenesis of Neonatal EncephalopathyDaniela MuñozNo ratings yet
- OligohydramniosDocument4 pagesOligohydramniossalamredNo ratings yet
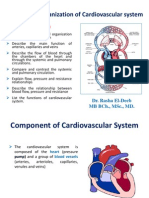
- Functional Organization of Cardiovascular SystemDocument19 pagesFunctional Organization of Cardiovascular SystemIbtesam Mohammed100% (2)
- Wiki Origin of The Domestic DogDocument38 pagesWiki Origin of The Domestic DogDe-hai CayolemNo ratings yet
- Bahasa InggrisDocument5 pagesBahasa Inggriskarina lestariNo ratings yet
- Laporan Kasus PPOKDocument23 pagesLaporan Kasus PPOKbahrinaNo ratings yet
- STRDocument179 pagesSTRJatin AnandNo ratings yet
- Natural Selection Lab-Phet Simulation Astrid 10DDocument8 pagesNatural Selection Lab-Phet Simulation Astrid 10DMary Fatima LlanesNo ratings yet
- WJMH 32 110Document6 pagesWJMH 32 110Yacine Tarik AizelNo ratings yet
- NLR Proteins - Methods and ProtocolsDocument269 pagesNLR Proteins - Methods and ProtocolsG DelisNo ratings yet
- Vaginal Examination Wit RationaleDocument1 pageVaginal Examination Wit RationaleAna Maria Teresa H RollanNo ratings yet
- Biological Basis of BehaviorDocument6 pagesBiological Basis of BehaviorLyanna MormontNo ratings yet
- Accepted Manuscript: Es Leblond, Emilie GayDocument45 pagesAccepted Manuscript: Es Leblond, Emilie GayCristhian Jover CastroNo ratings yet
- Labreportnew PDFDocument8 pagesLabreportnew PDFLuCky100% (1)
- United States v. PanlilioDocument4 pagesUnited States v. PanlilioZepht BadillaNo ratings yet
- HerpesDocument2 pagesHerpesSyafruddin GBNo ratings yet
- Types of NeuronsDocument10 pagesTypes of NeuronsJaspherBalderaNo ratings yet
- Revise Anatomy in 15 Days K Raviraj VD AgrawalDocument7 pagesRevise Anatomy in 15 Days K Raviraj VD AgrawalShehnaaz Khan29% (14)
- Zoo Season 1 EpisodesDocument4 pagesZoo Season 1 EpisodesValen CookNo ratings yet
- Mahasiswa STIKES RS. Baptis KediriDocument8 pagesMahasiswa STIKES RS. Baptis Kedirigia hisbian ladzianiNo ratings yet
- Respiratory Medicine 1 50Document33 pagesRespiratory Medicine 1 50Ahmed Kh. Abu WardaNo ratings yet
- Bites Stings and PoisonDocument78 pagesBites Stings and PoisonkagaralNo ratings yet