You might also like
- Microbial PhysiologyFrom EverandMicrobial PhysiologyAlbert G. MoatNo ratings yet
- NIH Public Access: Author ManuscriptDocument17 pagesNIH Public Access: Author ManuscriptSanthiya KunasegaranNo ratings yet
- 2017 Yasuda A Look at Plant Immunity Through The Window of The Multitasking Coreceptor BAK1Document9 pages2017 Yasuda A Look at Plant Immunity Through The Window of The Multitasking Coreceptor BAK1Raul DavilaNo ratings yet
- E2F-like Elements in p27 Promoter Specifically Sense Deregulated E2F ActivityDocument11 pagesE2F-like Elements in p27 Promoter Specifically Sense Deregulated E2F ActivityBell Villegas VNo ratings yet
- 1 s2.0 S1874939911001428 MainDocument11 pages1 s2.0 S1874939911001428 MainIzeeMohammadNo ratings yet
- Subfunctionalization of Peroxisome Proliferator Response Elements Accounts For Retention of Duplicated Fabp1 Genes in ZebrafishDocument15 pagesSubfunctionalization of Peroxisome Proliferator Response Elements Accounts For Retention of Duplicated Fabp1 Genes in ZebrafishTiago TorresNo ratings yet
- Differential Regulation of The Duplicated Fabp7, Fabp10 and Fabp11 Genes of Zebrafish by Peroxisome Proliferator Activated ReceptorsDocument10 pagesDifferential Regulation of The Duplicated Fabp7, Fabp10 and Fabp11 Genes of Zebrafish by Peroxisome Proliferator Activated ReceptorsTiago TorresNo ratings yet
- E2f1, A Novel Regulator of Metabolism: Introduction: A Cell Cycle Protein With New SkillsDocument8 pagesE2f1, A Novel Regulator of Metabolism: Introduction: A Cell Cycle Protein With New SkillsFaisal Fikri HakimNo ratings yet
- Microbe-Associated Molecular Patterns (Mamps) Probe Plant ImmunityDocument7 pagesMicrobe-Associated Molecular Patterns (Mamps) Probe Plant ImmunityJose Miguel Soto HerediaNo ratings yet
- Abdul Rafay Javed Roll No 23Document9 pagesAbdul Rafay Javed Roll No 23Rafay MirzaNo ratings yet
- 1 s2.0 S136952661400106X AmDocument20 pages1 s2.0 S136952661400106X AmElse12No ratings yet
- The Rb/E2F Pathway and Cancer: Joseph R. NevinsDocument6 pagesThe Rb/E2F Pathway and Cancer: Joseph R. NevinsSanthiya KunasegaranNo ratings yet
- Regulation of Purine Nucleotide Biosynthesis: in Yeast and BeyondDocument5 pagesRegulation of Purine Nucleotide Biosynthesis: in Yeast and Beyondsharna_kaydNo ratings yet
- Andrew Hu, 2018Document17 pagesAndrew Hu, 2018franciscaNo ratings yet
- ADAMs - Ohtsu Et Al - AJP Cell - 2006Document10 pagesADAMs - Ohtsu Et Al - AJP Cell - 2006Bilakovics NoemiNo ratings yet
- Ubiquitin ConjugationDocument9 pagesUbiquitin ConjugationDessiree PinkcessNo ratings yet
- BMC Evolutionary BiologyDocument11 pagesBMC Evolutionary Biologyannisa pramestiNo ratings yet
- Mauo 2018 Infeksi Ebv 2Document44 pagesMauo 2018 Infeksi Ebv 2OmeNo ratings yet
- ERADDocument7 pagesERADLary BagsNo ratings yet
- TMP 37 F4Document10 pagesTMP 37 F4FrontiersNo ratings yet
- 2011 Lopez-Gomez Interplay of Flg22-Induced Defence Responses and Nodulation in Lotus JaponicusDocument9 pages2011 Lopez-Gomez Interplay of Flg22-Induced Defence Responses and Nodulation in Lotus JaponicusRaul DavilaNo ratings yet
- New Twists in The Unfolded Protein Response: Cell BiologyDocument4 pagesNew Twists in The Unfolded Protein Response: Cell Biologyian8184No ratings yet
- Plasmodium Falciparum RIFIN Is A Novel Ligand For Inhibitory Immune Receptor LILRB2Document7 pagesPlasmodium Falciparum RIFIN Is A Novel Ligand For Inhibitory Immune Receptor LILRB2AlexTsuiNo ratings yet
- PP3 - 2009 - Dorsch - Nature Methods - Analysis of Receptor Oligomerization by FRAP MicrosDocument6 pagesPP3 - 2009 - Dorsch - Nature Methods - Analysis of Receptor Oligomerization by FRAP Microsdamon tanNo ratings yet
- Oligopeptide Permease in Borrelia Burgdorferi: Putative Peptide-Binding Components Encoded by Both Chromosomal and Plasmid LociDocument13 pagesOligopeptide Permease in Borrelia Burgdorferi: Putative Peptide-Binding Components Encoded by Both Chromosomal and Plasmid LocitkgehguoeNo ratings yet
- Luo 2016Document6 pagesLuo 2016Jose Alberto PbNo ratings yet
- Activacion of Plant Immune Responses by A Gain-Of-function Mutation in An Atypical Receptor-Like Kinase-Adolfo Jeueves 23 de Septiembre Del 2010Document9 pagesActivacion of Plant Immune Responses by A Gain-Of-function Mutation in An Atypical Receptor-Like Kinase-Adolfo Jeueves 23 de Septiembre Del 2010Laura Noriega CalixtoNo ratings yet
- Characterization of Arabidopsis Mutants That Are Associated With Altered C18 Unsaturated Fatty Acid MetabolismDocument7 pagesCharacterization of Arabidopsis Mutants That Are Associated With Altered C18 Unsaturated Fatty Acid MetabolismCamilo GaleanoNo ratings yet
- A J-Protein Co-Chaperone Recruits BiP To Monomerize IRE1 and Repress The Unfolded Protein ResponseDocument27 pagesA J-Protein Co-Chaperone Recruits BiP To Monomerize IRE1 and Repress The Unfolded Protein ResponseLefan YuNo ratings yet
- Research ProposalDocument7 pagesResearch ProposalBitan BiswasNo ratings yet
- Cell Signaling by Receptor Review Tyrosine Kinases: Joseph SchlessingerDocument15 pagesCell Signaling by Receptor Review Tyrosine Kinases: Joseph Schlessingergina inNo ratings yet
- PIIS1535947620300542Document15 pagesPIIS1535947620300542Blink SNo ratings yet
- Mass-spectrometric and bioinformatic analysis of eEF1Bγ interactome in the cytoplasmic fraction of A549 cellsDocument11 pagesMass-spectrometric and bioinformatic analysis of eEF1Bγ interactome in the cytoplasmic fraction of A549 cellsАнна ШаповаловаNo ratings yet
- TMP 34 B4Document4 pagesTMP 34 B4FrontiersNo ratings yet
- The Unfolded Protein Response - An OverviewDocument10 pagesThe Unfolded Protein Response - An OverviewgabrielpoulsonNo ratings yet
- Cloning, Purification and Comparative Characterization of Two Digestive Lysozymes From Musca Domestica LarvaeDocument9 pagesCloning, Purification and Comparative Characterization of Two Digestive Lysozymes From Musca Domestica LarvaebobyjuniorNo ratings yet
- Transforming Growth Factor Beta TGFBDocument8 pagesTransforming Growth Factor Beta TGFBxxxxxxxNo ratings yet
- UPR and VirusDocument26 pagesUPR and VirusAlex RomeroNo ratings yet
- TMP CEF6Document7 pagesTMP CEF6FrontiersNo ratings yet
- Red Cell: Blood GlycophorinsDocument11 pagesRed Cell: Blood GlycophorinsChu NgọcNo ratings yet
- Intracellular Proteins Involved in Inflammation and ApoptosisDocument12 pagesIntracellular Proteins Involved in Inflammation and ApoptosisJUAREZ GONZALEZ EDWARD HASSANNo ratings yet
- The Retinoblastoma Protein and Cell Cycle ControlDocument8 pagesThe Retinoblastoma Protein and Cell Cycle Controlaredhel ar-feinielNo ratings yet
- Types of Signalling Pathway Egfr Pathway Normal and DysregulatedDocument69 pagesTypes of Signalling Pathway Egfr Pathway Normal and Dysregulatedapi-251915360No ratings yet
- Inherited lrp2 Dysfunction in Human Disease and Animal ModelsDocument10 pagesInherited lrp2 Dysfunction in Human Disease and Animal Modelshennysusanto18No ratings yet
- TG Oe CEBPB OstopeniaDocument25 pagesTG Oe CEBPB OstopeniaNacido para BendcirNo ratings yet
- L Dopa VitreoscillaDocument12 pagesL Dopa VitreoscillaJesús SaenzNo ratings yet
- Jurnal2 PDFDocument12 pagesJurnal2 PDFRINDA HERNISNo ratings yet
- HY5 Protein in ArabidopsisDocument11 pagesHY5 Protein in ArabidopsismNo ratings yet
- PIIS0960982214005296Document8 pagesPIIS0960982214005296Sofia Oliveira SantosNo ratings yet
- Raghavendra Et Al (2010) ABA Di Lahan KekeringanDocument7 pagesRaghavendra Et Al (2010) ABA Di Lahan KekeringanizdamaitaNo ratings yet
- Ergo Va LineDocument9 pagesErgo Va LineMax SalesNo ratings yet
- The Major Human Structural IgE Epitope of The Brazil Nut Allergen Ber e 1 - A Chimaeric and Protein Microarray ApproachDocument11 pagesThe Major Human Structural IgE Epitope of The Brazil Nut Allergen Ber e 1 - A Chimaeric and Protein Microarray ApproachponbohacopNo ratings yet
- Intl Journal of Cancer - 2015 - Inoue - Transcription Factors That Interact With p53 and Mdm2Document9 pagesIntl Journal of Cancer - 2015 - Inoue - Transcription Factors That Interact With p53 and Mdm2JESUS DAVID VALAREZO QUEVEDONo ratings yet
- Cabaton I In. 2009Document10 pagesCabaton I In. 2009Ola ZygmuntowiczNo ratings yet
- Buonanno 2001Document10 pagesBuonanno 2001Maurício DallowayNo ratings yet
- Caunt 2015Document16 pagesCaunt 2015Thanasis PapatheodorouNo ratings yet
- Christian Hiepen Et AlDocument47 pagesChristian Hiepen Et AlJOSE LUIS CUELLAR CAMACHONo ratings yet
- Anti Cancer Activity Prediction of Secondary Metabolites From Marine Sponge Discodermia Calyx: An in Silico ApproachDocument6 pagesAnti Cancer Activity Prediction of Secondary Metabolites From Marine Sponge Discodermia Calyx: An in Silico ApproachNovayani PagilingNo ratings yet
- Influence of FOX genes on aging and aging-associated diseasesFrom EverandInfluence of FOX genes on aging and aging-associated diseasesNo ratings yet
- Andrasi 2019 JEBDocument15 pagesAndrasi 2019 JEBAladár Pettkó-SzandtnerNo ratings yet
- Letoha 2019 SciRepDocument17 pagesLetoha 2019 SciRepAladár Pettkó-SzandtnerNo ratings yet
- Dicot and Monocot Plants Differ in Retinoblastoma-Related Protein SubfamiliesDocument13 pagesDicot and Monocot Plants Differ in Retinoblastoma-Related Protein SubfamiliesAladár Pettkó-SzandtnerNo ratings yet
- Elife-29845-V1 Hsp70-Associated Chaperones Have A Critical Role in Buffering Protein Production CostsDocument23 pagesElife-29845-V1 Hsp70-Associated Chaperones Have A Critical Role in Buffering Protein Production CostsAladár Pettkó-SzandtnerNo ratings yet
- Dev170639.full Drosophila Small Ovary Gene Is Required For Transposon Silencing and Heterochromatin Organization, and Ensures Germline Stem Cell Maintenance and DifferentiationDocument13 pagesDev170639.full Drosophila Small Ovary Gene Is Required For Transposon Silencing and Heterochromatin Organization, and Ensures Germline Stem Cell Maintenance and DifferentiationAladár Pettkó-SzandtnerNo ratings yet
- Elife-29845-V1 Hsp70-Associated Chaperones Have A Critical Role in Buffering Protein Production CostsDocument23 pagesElife-29845-V1 Hsp70-Associated Chaperones Have A Critical Role in Buffering Protein Production CostsAladár Pettkó-SzandtnerNo ratings yet
- Core Cell Cycle Regulatory Genes in Rice and Their Expression Profiles Across The Growth Zone of The LeafDocument41 pagesCore Cell Cycle Regulatory Genes in Rice and Their Expression Profiles Across The Growth Zone of The LeafAladár Pettkó-SzandtnerNo ratings yet
- You Have Access E2FB Interacts With RETINOBLASTOMA RELATED and Regulates Cell Proliferation During Leaf DevelopmentDocument49 pagesYou Have Access E2FB Interacts With RETINOBLASTOMA RELATED and Regulates Cell Proliferation During Leaf DevelopmentAladár Pettkó-SzandtnerNo ratings yet
- ResponseToReviewers Cserhati RiceCC JPR FinalDocument4 pagesResponseToReviewers Cserhati RiceCC JPR FinalAladár Pettkó-SzandtnerNo ratings yet
- Pettko-Szandtner Et AlDocument13 pagesPettko-Szandtner Et AlAladár Pettkó-SzandtnerNo ratings yet
- Advia Centaur SafetyDocument5 pagesAdvia Centaur SafetyMaduraiveeran cNo ratings yet
- CBST LastDocument5 pagesCBST LastMozma AwanNo ratings yet
- ECGC PO Descriptive BookDocument74 pagesECGC PO Descriptive BookAmar Pandey100% (2)
- Adams4e Tif Ch47Document19 pagesAdams4e Tif Ch47fbernis1480_11022046100% (1)
- Lung Cancer Prediction Using Data Mining TechniquesDocument6 pagesLung Cancer Prediction Using Data Mining TechniquesKEZZIA MAE ABELLANo ratings yet
- Veterinary Internal Medicne - 2008 - Bruchim - Heat Stroke in Dogs A Retrospective Study of 54 Cases 1999 2004 andDocument9 pagesVeterinary Internal Medicne - 2008 - Bruchim - Heat Stroke in Dogs A Retrospective Study of 54 Cases 1999 2004 andGuillermo MuzasNo ratings yet
- 3.ALS Algorithms Advanced Life SupportDocument1 page3.ALS Algorithms Advanced Life SupportLucian Alin DinuNo ratings yet
- Assessment Dianosis Planning Intervention Rationale EvaluationDocument2 pagesAssessment Dianosis Planning Intervention Rationale Evaluationkyaw100% (1)
- Respiratory PhysiologyDocument16 pagesRespiratory PhysiologyYsabel Salvador DychincoNo ratings yet
- Conversion of ASTM To TBP and EFVDocument111 pagesConversion of ASTM To TBP and EFVsyedmuhammadtariqueNo ratings yet
- Arcega ErlindaDocument7 pagesArcega ErlindaIvonne TernidaNo ratings yet
- New Male StudiesDocument12 pagesNew Male StudiesPaul Andrey DichosonNo ratings yet
- # Category Image Product Name Producer Package Hs Code Remark Key FeaturesDocument9 pages# Category Image Product Name Producer Package Hs Code Remark Key Featuresiq_dianaNo ratings yet
- 9 Steps To Reverse DementiaDocument36 pages9 Steps To Reverse DementiaLavinia PirlogNo ratings yet
- Low Cost Thermal Camera For Use in Preclinical Detection of Diabetic PeripheralDocument8 pagesLow Cost Thermal Camera For Use in Preclinical Detection of Diabetic PeripheralEDGAR ISMAEL REYES DEZANo ratings yet
- Cognitive Impairment Along The Course of Depression: Non-Pharmacological Treatment OptionsDocument11 pagesCognitive Impairment Along The Course of Depression: Non-Pharmacological Treatment OptionsCarolina PradoNo ratings yet
- Anaphy and Patho of AppendicitisDocument3 pagesAnaphy and Patho of AppendicitisArt Christian RamosNo ratings yet
- Common Questions About Oppositional Defiant Disorder - American Family PhysicianDocument12 pagesCommon Questions About Oppositional Defiant Disorder - American Family Physiciando lee100% (1)
- The Nurse Leader's Role: Crisis ManagementDocument3 pagesThe Nurse Leader's Role: Crisis Managementravsab GaikwadNo ratings yet
- Armohib 28 MSDSDocument8 pagesArmohib 28 MSDSdalton2004No ratings yet
- Icd 10Document54 pagesIcd 10gexchaNo ratings yet
- PCT For BodybuildersDocument12 pagesPCT For BodybuildersJon Mester100% (4)
- Ultimate Guide To Surviving in The WildDocument175 pagesUltimate Guide To Surviving in The WildSal Ot100% (1)
- DISEASE DETECTION IN VEGETABLES (TOMATO) USING DEEP LEARNINgDocument38 pagesDISEASE DETECTION IN VEGETABLES (TOMATO) USING DEEP LEARNINgA ANo ratings yet
- 5.RTRI Principle and Performance 8may2019Document34 pages5.RTRI Principle and Performance 8may2019ABCDeNo ratings yet
- Full Download Test Bank For Essentials of Genetics 8th Edition by Klug PDF Full ChapterDocument36 pagesFull Download Test Bank For Essentials of Genetics 8th Edition by Klug PDF Full Chapterfencingvesper9dgb04100% (17)
- Teaching Medical Students Basic Principles of Laboratory MedicineDocument14 pagesTeaching Medical Students Basic Principles of Laboratory MedicineIrish De VeraNo ratings yet
- Statistics Notes in The British Medical Journal (Bland JM, Altman DG. - NEJ)Document95 pagesStatistics Notes in The British Medical Journal (Bland JM, Altman DG. - NEJ)pegazus_arNo ratings yet
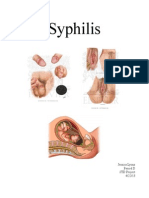
- Syphilis: Jessica Lyons Period D STD Project 4/22/13Document3 pagesSyphilis: Jessica Lyons Period D STD Project 4/22/13jesssielynnn33No ratings yet
- Concept PaperDocument14 pagesConcept PaperRyan Maghanoy100% (3)