You might also like
- Cloning and Characterization of A Water Deficit Stress Responsive Transcription Factor Gene From Oryza Sativa LDocument11 pagesCloning and Characterization of A Water Deficit Stress Responsive Transcription Factor Gene From Oryza Sativa Lenoch taclanNo ratings yet
- Ribosome-inactivating Proteins: Ricin and Related ProteinsFrom EverandRibosome-inactivating Proteins: Ricin and Related ProteinsFiorenzo StirpeNo ratings yet
- Raphanus Sativus (Radish) : Their Chemistry: and BiologyDocument28 pagesRaphanus Sativus (Radish) : Their Chemistry: and BiologyAhmed Gh Al-zechrawiNo ratings yet
- Garuda 1228838Document7 pagesGaruda 1228838hadi.wicaksanaNo ratings yet
- Isolation of Two Edna Clones Coding For Larval Hemolymph Proteins ofDocument15 pagesIsolation of Two Edna Clones Coding For Larval Hemolymph Proteins ofplatusNo ratings yet
- J5fhhghgbb776ohhh7810urnal Pooone 0219973Document18 pagesJ5fhhghgbb776ohhh7810urnal Pooone 0219973Red DiggerNo ratings yet
- Carbohydrate Research: Werner Praznik, Renate Löppert, Josè M. Cruz Rubio, Klaus Zangger, Anton HuberDocument10 pagesCarbohydrate Research: Werner Praznik, Renate Löppert, Josè M. Cruz Rubio, Klaus Zangger, Anton HuberDiane FauziNo ratings yet
- Chen 2016Document9 pagesChen 2016Shampa SenNo ratings yet
- tmp7F3E TMPDocument6 pagestmp7F3E TMPFrontiersNo ratings yet
- Styer Et - Al - 2004Document7 pagesStyer Et - Al - 2004ieremiasNo ratings yet
- Cicer ArietinumDocument7 pagesCicer ArietinumSaul Dominguez SmithNo ratings yet
- Cloning, Purification and Comparative Characterization of Two Digestive Lysozymes From Musca Domestica LarvaeDocument9 pagesCloning, Purification and Comparative Characterization of Two Digestive Lysozymes From Musca Domestica LarvaebobyjuniorNo ratings yet
- Lactobacillus SPP.: Lactobacillus Plantarum: Taxonomy, Identification, and Genetics of The SpeciesDocument8 pagesLactobacillus SPP.: Lactobacillus Plantarum: Taxonomy, Identification, and Genetics of The SpeciesAlex CárdenasNo ratings yet
- PGR Arun 2011 BDocument5 pagesPGR Arun 2011 Barunprabhu_dhanapalNo ratings yet
- Stas Olla 2003Document25 pagesStas Olla 2003lizbethNo ratings yet
- Escherichia Coli Appa Encoded BifunctionalDocument13 pagesEscherichia Coli Appa Encoded BifunctionalMarco RamosNo ratings yet
- 1989-08 Cationic Liposome-Mdeiated RNA Transfection-6077.fullDocument5 pages1989-08 Cationic Liposome-Mdeiated RNA Transfection-6077.fullRetro ActiveNo ratings yet
- Enzymatic Total Synthesis of Gibberellin A4Document8 pagesEnzymatic Total Synthesis of Gibberellin A4ranukahewageNo ratings yet
- 35IJEAB 102202112 EffectofDocument8 pages35IJEAB 102202112 EffectofIJEAB JournalNo ratings yet
- Functional Expression of Horseradish Peroxidase in Saccharomyces Cerevisiae and Pichia PastorisDocument8 pagesFunctional Expression of Horseradish Peroxidase in Saccharomyces Cerevisiae and Pichia PastorisAlan SublimeNo ratings yet
- Ergo Va LineDocument9 pagesErgo Va LineMax SalesNo ratings yet
- 1-s2.0-S0378874122006729-mainDocument16 pages1-s2.0-S0378874122006729-mainSubhadip BanerjeeNo ratings yet
- Preparation of Kavalactone PrecursorDocument9 pagesPreparation of Kavalactone PrecursorEchobackNo ratings yet
- Isolation and Expression Analysis of A GDSL-like Lipase Gene From LDocument7 pagesIsolation and Expression Analysis of A GDSL-like Lipase Gene From LBudi Hermanto MadjagaNo ratings yet
- Arabidopsis Thaliana Has The Catalytic Capacity To ActivateDocument11 pagesArabidopsis Thaliana Has The Catalytic Capacity To ActivateFrontiersNo ratings yet
- 10 1002@bies 20085Document12 pages10 1002@bies 20085Minh Nguyen NgocNo ratings yet
- Lipid Metabolism in The Larval and Adult Forms ofDocument9 pagesLipid Metabolism in The Larval and Adult Forms ofdonsoucy1069No ratings yet
- CarbohydratDocument6 pagesCarbohydratVan Anh NguyenNo ratings yet
- 16S Ribosomal DNA For: Amplification Phylogenetic StudyDocument7 pages16S Ribosomal DNA For: Amplification Phylogenetic StudyolamicroNo ratings yet
- The Substrate Specificity-Determining Amino Acid Code of 4-Coumarate:Coa LigaseDocument6 pagesThe Substrate Specificity-Determining Amino Acid Code of 4-Coumarate:Coa LigaseFrontiersNo ratings yet
- TMP 7 E9 EDocument13 pagesTMP 7 E9 EFrontiersNo ratings yet
- Gmdreb2, A Soybean Dre-Binding Transcription Factor, Conferred Drought and High-Salt Tolerance in Transgenic PlantsDocument7 pagesGmdreb2, A Soybean Dre-Binding Transcription Factor, Conferred Drought and High-Salt Tolerance in Transgenic PlantsOscar VanegasNo ratings yet
- Differential Expression ... Unicellular ChlorophyteDocument7 pagesDifferential Expression ... Unicellular ChlorophyteAJITHKUMARNo ratings yet
- Molecules 24 02408Document13 pagesMolecules 24 02408Sudhir Kumar MishraNo ratings yet
- Latex Clearing Protein-An Oxygenase Cleaving Poly (Cis-1,4-Isoprene) Rubber at The Cis Double BondsDocument10 pagesLatex Clearing Protein-An Oxygenase Cleaving Poly (Cis-1,4-Isoprene) Rubber at The Cis Double BondsCristian ValdésNo ratings yet
- Differential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsisDocument13 pagesDifferential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsismNo ratings yet
- Ramadan2021 Article TheFirstReportOfRNAUToCOrGEditDocument8 pagesRamadan2021 Article TheFirstReportOfRNAUToCOrGEditFika Ayu SafitriNo ratings yet
- CRISPR Cas9Document9 pagesCRISPR Cas9Vladislav TimoshenkoNo ratings yet
- Am18 Program WDocument8 pagesAm18 Program WHan-Nam PHAMNo ratings yet
- Analysis of Food Oligosaccharides Using MALDI-MS: Quantification of FructooligosaccharidesDocument9 pagesAnalysis of Food Oligosaccharides Using MALDI-MS: Quantification of FructooligosaccharidesseverithNo ratings yet
- PavllovaDocument5 pagesPavllovalucia_bregNo ratings yet
- GCE A Level Biology ExamDocument6 pagesGCE A Level Biology ExamjoannetzyNo ratings yet
- Yoshida 2006 FA MolDocument12 pagesYoshida 2006 FA MolBruno de Moraes NunesNo ratings yet
- JBC 2002 FBA Vs TBADocument7 pagesJBC 2002 FBA Vs TBAVincent GeruszNo ratings yet
- WebOfScience-5736Document1,724 pagesWebOfScience-5736Hoàng Duy ĐỗNo ratings yet
- J. Biol. Chem.-1984-Place-1299-308Document11 pagesJ. Biol. Chem.-1984-Place-1299-308João AlvesNo ratings yet
- AVOCADO CANCER PATHWAYDocument5 pagesAVOCADO CANCER PATHWAYfelipe2000ipnNo ratings yet
- 1197-Article Text-2669-1-10-20211031Document5 pages1197-Article Text-2669-1-10-20211031Kiệt Phan AnhNo ratings yet
- (Glossonema Edule) : Constituents O F Plants Growing in Qatar Iiit - Nutritive Constituents O F GarawahDocument6 pages(Glossonema Edule) : Constituents O F Plants Growing in Qatar Iiit - Nutritive Constituents O F GarawahJuan Manuel C FigueroaNo ratings yet
- Tugas JurnalDocument11 pagesTugas Jurnalsherly handayaniNo ratings yet
- Metabolic Engineering of Carotenoid Accumulation in E.coliDocument5 pagesMetabolic Engineering of Carotenoid Accumulation in E.coliAJITHKUMARNo ratings yet
- 2015-A Japanese Plum Alpha-L-Arabinofuranosidase - Beta-D-Xylosidase Gene Is Developmentally Regulated by Alternative SplicingDocument11 pages2015-A Japanese Plum Alpha-L-Arabinofuranosidase - Beta-D-Xylosidase Gene Is Developmentally Regulated by Alternative SplicingEduardo PaganoNo ratings yet
- Zheng 2005Document12 pagesZheng 2005Bartolo Concha FríasNo ratings yet
- Plant Carotinoids BiochemistryDocument14 pagesPlant Carotinoids Biochemistrypaul WongNo ratings yet
- Gene sequence and protein analysis of yeast MEL1 gene encoding secreted alpha-galactosidaseDocument8 pagesGene sequence and protein analysis of yeast MEL1 gene encoding secreted alpha-galactosidaseVijendra KavatalkarNo ratings yet
- Rapid Changes in Elongation Factor Transcripts During Sea Urchin DevelopmentDocument3 pagesRapid Changes in Elongation Factor Transcripts During Sea Urchin DevelopmentNada ZnaidiNo ratings yet
- Parp 1 e 2Document6 pagesParp 1 e 2AnnalisaDorioNo ratings yet
- PopuSSR - Primer Lation Structure Analysis 1.2Document8 pagesPopuSSR - Primer Lation Structure Analysis 1.2RITESH CHANDANo ratings yet
- BMB 117Document12 pagesBMB 117Ironic SunNo ratings yet
- Dwnload Full Psychology Core Concepts 8th Edition Zimbardo Solutions Manual PDFDocument32 pagesDwnload Full Psychology Core Concepts 8th Edition Zimbardo Solutions Manual PDFcomfortdehm1350100% (8)
- O M Study Guide AnswerkeyDocument2 pagesO M Study Guide Answerkeyapi-237676607No ratings yet
- KCSE PREMOCKS Set 2Document262 pagesKCSE PREMOCKS Set 2Micah IsabokeNo ratings yet
- 2016 HGVS Recommendations For The Description of SequenceVariants (2016 Update)Document6 pages2016 HGVS Recommendations For The Description of SequenceVariants (2016 Update)Alfonso PeñarroyaNo ratings yet
- AP Biology Chapter 14 - Mendel and The Gene IdeaDocument3 pagesAP Biology Chapter 14 - Mendel and The Gene IdeawaterforlifeNo ratings yet
- Honors Biology: 21-22 Midterm Exam Review PacketDocument11 pagesHonors Biology: 21-22 Midterm Exam Review PacketaubreymertNo ratings yet
- Botany OvuleDocument5 pagesBotany OvuleMisbah IqbalNo ratings yet
- Introduction To Cyanophyta: Cell Structures ContDocument4 pagesIntroduction To Cyanophyta: Cell Structures ContClaudia Yuli AstutieNo ratings yet
- Shao 2019Document8 pagesShao 2019Muhamad Akbar NamusyNo ratings yet
- Explain The Interpretation of Hemoglobin Result (Low Indicator Result)Document5 pagesExplain The Interpretation of Hemoglobin Result (Low Indicator Result)Gede NgurahNo ratings yet
- Hickman Zoology 18e PDFDocument4 pagesHickman Zoology 18e PDFSalsabila Tasyari0% (1)
- Unit 4 Study GuideDocument3 pagesUnit 4 Study Guidesawyeranthony2009No ratings yet
- Microbes and Infection: Jin-Yan Li, Zhi You, Qiong Wang, Zhi-Jian Zhou, Ye Qiu, Rui Luo, Xing-Yi GeDocument6 pagesMicrobes and Infection: Jin-Yan Li, Zhi You, Qiong Wang, Zhi-Jian Zhou, Ye Qiu, Rui Luo, Xing-Yi GeLeah StroposNo ratings yet
- Daftar Pustaka - 1Document4 pagesDaftar Pustaka - 1Rama YeniNo ratings yet
- 01 01aLectureNotesDocument16 pages01 01aLectureNotesRosalie Jane OrtizNo ratings yet
- Yeast DisplayDocument7 pagesYeast DisplayAnnisa Pratiwi GunawanNo ratings yet
- N Comms 14644Document16 pagesN Comms 14644Vivien JusztusNo ratings yet
- All Animals A-Z List - Animal Names - AZ AnimalsDocument2 pagesAll Animals A-Z List - Animal Names - AZ AnimalsArch. ZionNo ratings yet
- Gen WorksheetDocument2 pagesGen WorksheetVi HuynhNo ratings yet
- Introduction to EEG: An Overview of Biomedical Signal Processing ConceptsDocument25 pagesIntroduction to EEG: An Overview of Biomedical Signal Processing ConceptsImesh Prathiba Lihinikaduarachchi100% (1)
- Controlling Factors in Craniofacial GrowthDocument82 pagesControlling Factors in Craniofacial GrowthPrachodh0% (1)
- Gen BankDocument6 pagesGen Bankmcdonald212No ratings yet
- Cell Cycle RegulationDocument2 pagesCell Cycle RegulationAhmed FazlyNo ratings yet
- Respiration Lec10 SlidesDocument29 pagesRespiration Lec10 SlidesHussain IbrahimNo ratings yet
- Microorganisms Test Multiple ChoiceDocument5 pagesMicroorganisms Test Multiple ChoiceBalan MuthuNo ratings yet
- 3 - IB Biology (2016) - 1.3 - Membrane Structure PPT Part BDocument10 pages3 - IB Biology (2016) - 1.3 - Membrane Structure PPT Part BUlises DeyesNo ratings yet
- Allgoewer Et Al-2001-Veterinary OphthalmologyDocument6 pagesAllgoewer Et Al-2001-Veterinary OphthalmologyRose CavalcanteNo ratings yet
- GATE Life SciencesDocument6 pagesGATE Life SciencesstudyurselfNo ratings yet
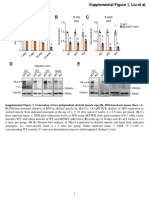
- MLL4 Supplemental Figure 136155-JCI-RG-RV-3 - SD - 424851Document15 pagesMLL4 Supplemental Figure 136155-JCI-RG-RV-3 - SD - 424851李博No ratings yet
- Lecture 2Document32 pagesLecture 2alizeh amanNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationFrom EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationRating: 4 out of 5 stars4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- Fearfully and Wonderfully: The Marvel of Bearing God's ImageFrom EverandFearfully and Wonderfully: The Marvel of Bearing God's ImageRating: 5 out of 5 stars5/5 (40)
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)