You might also like
- Statistics for Bioinformatics: Methods for Multiple Sequence AlignmentFrom EverandStatistics for Bioinformatics: Methods for Multiple Sequence AlignmentNo ratings yet
- Gahan 2013 Acidos Nucleicos Efectos InherenciaDocument18 pagesGahan 2013 Acidos Nucleicos Efectos InherenciaKaritoNo ratings yet
- Genetic Signatures of Evolution of The Pluripotency Gene Regulating Network Across MammalsDocument13 pagesGenetic Signatures of Evolution of The Pluripotency Gene Regulating Network Across MammalsSevim AkçağlarNo ratings yet
- Genome-Wide Alternative Polyadenylation in Animals: Insights From High-Throughput TechnologiesDocument10 pagesGenome-Wide Alternative Polyadenylation in Animals: Insights From High-Throughput TechnologiesdupuytrenNo ratings yet
- Infection, Genetics and Evolution: Haemaphysalis QinghaiensisDocument15 pagesInfection, Genetics and Evolution: Haemaphysalis QinghaiensisChickencito PerezNo ratings yet
- Exosome Biogenesis Bioactivities and Functions As New de 2018 BiotechnologyDocument7 pagesExosome Biogenesis Bioactivities and Functions As New de 2018 BiotechnologyMally JamesNo ratings yet
- Comp Transcript Between Zebrafsh and MammalsDocument5 pagesComp Transcript Between Zebrafsh and MammalsRita RodriguesNo ratings yet
- Fpls 05 00209 1.pdf - SosDocument15 pagesFpls 05 00209 1.pdf - SosCHATZIKYRIAKIDOUNo ratings yet
- 1 s2.0 S0168165623000123 MainDocument10 pages1 s2.0 S0168165623000123 MainLauren Caluya LañoNo ratings yet
- tmp2739 TMPDocument3 pagestmp2739 TMPFrontiersNo ratings yet
- Fgene 11 00995Document12 pagesFgene 11 009954jr8tkzs7dNo ratings yet
- 10.1007@s11427 020 1709 6Document4 pages10.1007@s11427 020 1709 6Nouri RaidNo ratings yet
- Group 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroDocument4 pagesGroup 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroAnne Olfato ParojinogNo ratings yet
- 2019 Cao Lipshitz BiorxivDocument39 pages2019 Cao Lipshitz BiorxivAndrew LiebermannNo ratings yet
- latenciaDocument12 pageslatenciageorgedNo ratings yet
- 1-s2.0-S1749461317300611-mainDocument5 pages1-s2.0-S1749461317300611-mainBen DresimNo ratings yet
- Zhao Et Al 2021 Pentatoma Rufipes MitogenomeDocument22 pagesZhao Et Al 2021 Pentatoma Rufipes MitogenomeDávid RédeiNo ratings yet
- Guo Et Al. 2008Document11 pagesGuo Et Al. 2008Jose FernandoNo ratings yet
- Hiergeist Et Al, 2015 - Analise Da Microbiota Intestinal HumanoDocument13 pagesHiergeist Et Al, 2015 - Analise Da Microbiota Intestinal HumanoanavcmeloNo ratings yet
- Bioinformatical approaches to investigating transcription factor binding sitesDocument5 pagesBioinformatical approaches to investigating transcription factor binding sitesAiman RoslizarNo ratings yet
- Short Tandem Repeats (STR) in Cattle Genomics and BreedingDocument21 pagesShort Tandem Repeats (STR) in Cattle Genomics and BreedingDušica Ostojić AndrićNo ratings yet
- Oven Et Al-2009-Human MutationDocument9 pagesOven Et Al-2009-Human MutationabidaiqbalansariNo ratings yet
- 581 FullDocument10 pages581 FullMhemeydha Luphe YudhaNo ratings yet
- (14796805 - Journal of Endocrinology) Mouse Models For Inherited Endocrine and Metabolic DisordersDocument20 pages(14796805 - Journal of Endocrinology) Mouse Models For Inherited Endocrine and Metabolic DisordersJohn ShipNo ratings yet
- KEY DATA FROM THE 2022 EUROPEAN THYROID ASSOCIATION CONGRESS. Role of Molecular Biology in Aggressive Thyroid CancerDocument13 pagesKEY DATA FROM THE 2022 EUROPEAN THYROID ASSOCIATION CONGRESS. Role of Molecular Biology in Aggressive Thyroid Cancermiguel saba sabaNo ratings yet
- Articulo 3Document3 pagesArticulo 3u20211196389No ratings yet
- Female and Male Gamete Mitochondria Are Distinct and Complementary in Transcription, Structure, and Genome FunctionDocument9 pagesFemale and Male Gamete Mitochondria Are Distinct and Complementary in Transcription, Structure, and Genome FunctionardirshajiNo ratings yet
- Functional Bias and Demographic History Obscure PaDocument46 pagesFunctional Bias and Demographic History Obscure PaGuada AguirreNo ratings yet
- Evolu IntronesDocument5 pagesEvolu IntronesIsaías Turrubiate PNo ratings yet
- Articulo 5Document12 pagesArticulo 5Holman SanabriaNo ratings yet
- Microsatellite Markers: What They Mean and Why They Are So UsefulDocument17 pagesMicrosatellite Markers: What They Mean and Why They Are So UsefulNTA UGC-NETNo ratings yet
- (17417899 - Reproduction) Mitochondrial Humanin Peptide Acts As A Cytoprotective Factor in Granulosa Cell SurvivalDocument11 pages(17417899 - Reproduction) Mitochondrial Humanin Peptide Acts As A Cytoprotective Factor in Granulosa Cell SurvivalLeilane GlienkeNo ratings yet
- Busulfan Induces Apoptotic and Cytotoxic Effects On Testis and Epididymal Sperm of Adult Male Mouse Following Low Dose TreatmentDocument9 pagesBusulfan Induces Apoptotic and Cytotoxic Effects On Testis and Epididymal Sperm of Adult Male Mouse Following Low Dose TreatmentInternational Network For Natural SciencesNo ratings yet
- Jkaa 022Document8 pagesJkaa 022李宜芸No ratings yet
- Genome Res.-2013-Alföldi-1063-8Document7 pagesGenome Res.-2013-Alföldi-1063-8Yamile A Rodríguez RiascosNo ratings yet
- Revista Brasileira de Zootecnia: Caiyun Fan, Yunhai Zhang, Juhua Wang, Jianbo ChengDocument8 pagesRevista Brasileira de Zootecnia: Caiyun Fan, Yunhai Zhang, Juhua Wang, Jianbo ChengyutefupNo ratings yet
- Check_rapposa_Highly Conserved D-loop Sequences in Woolly Mouse Opossums MarmosaDocument8 pagesCheck_rapposa_Highly Conserved D-loop Sequences in Woolly Mouse Opossums Marmosagbitencourt.bioNo ratings yet
- Molecular Cloning and Expression Analysis of The Ajuba Gene of Grass Carp (Ctenopharyngodon Idella) Involved in Cellular Response To Viral InfectionDocument7 pagesMolecular Cloning and Expression Analysis of The Ajuba Gene of Grass Carp (Ctenopharyngodon Idella) Involved in Cellular Response To Viral InfectionRizka Rahmana PutriNo ratings yet
- tmp564B TMPDocument3 pagestmp564B TMPFrontiersNo ratings yet
- Large-Scale Chemical-Genetics of The Human Gut Bacterium Bacteroides ThetaiotaomicronDocument35 pagesLarge-Scale Chemical-Genetics of The Human Gut Bacterium Bacteroides ThetaiotaomicronKarl SaganNo ratings yet
- Metástase IngDocument22 pagesMetástase IngAmanda SophiaNo ratings yet
- Msab 296Document15 pagesMsab 296Ches PascualNo ratings yet
- International Journal For ParasitologyDocument10 pagesInternational Journal For ParasitologyGabriel AvilaNo ratings yet
- Drosophila: Modeling Tumor Invasion and Metastasis inDocument9 pagesDrosophila: Modeling Tumor Invasion and Metastasis inFernando Lazzarotto EvaldtNo ratings yet
- Artigo 1 - Mitochondrial EvolutionDocument17 pagesArtigo 1 - Mitochondrial EvolutionMax NunesNo ratings yet
- Articulo 1Document6 pagesArticulo 1Fabio Andrés EnríquezNo ratings yet
- 1 s2.0 S0888754323000332 MainDocument44 pages1 s2.0 S0888754323000332 MainNour MhiriNo ratings yet
- Genome-Wide Analysis of The Arabidopsis Thaliana Replication Timing ProgramDocument66 pagesGenome-Wide Analysis of The Arabidopsis Thaliana Replication Timing ProgramZahid Yaqoob BhatNo ratings yet
- Transgenic Animals: (From CCAC Resource Supplement, Spring/Summer 1997. Used With Permission of CCAC)Document5 pagesTransgenic Animals: (From CCAC Resource Supplement, Spring/Summer 1997. Used With Permission of CCAC)nitralekhaNo ratings yet
- Science-2004-Leroy EbolaDocument6 pagesScience-2004-Leroy EbolaJuan MoralesNo ratings yet
- Neurological and Physiological Disorders in Artemia Harboring Manipulative CestodesDocument5 pagesNeurological and Physiological Disorders in Artemia Harboring Manipulative CestodeslacosNo ratings yet
- Architecture of The Outbred Brown Fat Proteome DefDocument49 pagesArchitecture of The Outbred Brown Fat Proteome Defh5wwgbvbvmNo ratings yet
- tmp9855 TMPDocument9 pagestmp9855 TMPFrontiersNo ratings yet
- Tnt1 Retrotransposon MutagenesisDocument12 pagesTnt1 Retrotransposon MutagenesisEliotKellyNo ratings yet
- 1-s2.0-S1749461320300257-mainDocument8 pages1-s2.0-S1749461320300257-mainBen DresimNo ratings yet
- The Architecture of A Scrambled Genome Reveals Massive Levels of Genomic Rearrangement During DevelopmentDocument12 pagesThe Architecture of A Scrambled Genome Reveals Massive Levels of Genomic Rearrangement During DevelopmentAnonymous slVH85zYNo ratings yet
- Mol Biol Evol 2011 Paar 1877 92Document16 pagesMol Biol Evol 2011 Paar 1877 92gunjan1920No ratings yet
- Systems Biology in Animal Production and Health, Volume 1Document161 pagesSystems Biology in Animal Production and Health, Volume 1CutlerBeckett90No ratings yet
- A Neuroscientist S Guide To Transgenic Mice An - 2020 - Neuroscience - BiobehaviDocument17 pagesA Neuroscientist S Guide To Transgenic Mice An - 2020 - Neuroscience - BiobehaviNabil Sangga BuanaNo ratings yet
- Genome Res.-2007-He-1471-7Document8 pagesGenome Res.-2007-He-1471-7Richard Icaro EsparzaNo ratings yet
- Edunaturalia: Analisis Pemahaman Siswa Sma Di Kota Pontianak Mengenai Materi GenetikaDocument6 pagesEdunaturalia: Analisis Pemahaman Siswa Sma Di Kota Pontianak Mengenai Materi Genetikaa lNo ratings yet
- Acad Dermatol Venereol - 2015 - Lambert - Real Life Effectiveness of Once Daily Calcipotriol and Betamethasone DipropionateDocument7 pagesAcad Dermatol Venereol - 2015 - Lambert - Real Life Effectiveness of Once Daily Calcipotriol and Betamethasone Dipropionatea lNo ratings yet
- Bioindicators The Natural Indicator of Environmental PollutionDocument10 pagesBioindicators The Natural Indicator of Environmental PollutionAngie GonzálezNo ratings yet
- Age, Gender, and Muscle Strength: A Study Based On Indonesian SamplesDocument7 pagesAge, Gender, and Muscle Strength: A Study Based On Indonesian Samplesa lNo ratings yet
- MitosisDocument7 pagesMitosisa lNo ratings yet
- MitosisDocument7 pagesMitosisa lNo ratings yet
- 1 s2.0 S0168169916303507 Main PDFDocument15 pages1 s2.0 S0168169916303507 Main PDFa lNo ratings yet
- Turning Meiosis Into MitosisDocument10 pagesTurning Meiosis Into Mitosisa lNo ratings yet
- Cytological Effects of Blitox On Root Mitosis of Allium: Cepa LDocument7 pagesCytological Effects of Blitox On Root Mitosis of Allium: Cepa La lNo ratings yet
- 1 s2.0 S0168169916303507 Main PDFDocument15 pages1 s2.0 S0168169916303507 Main PDFa lNo ratings yet
- 1 s2.0 S0168169916303507 Main PDFDocument15 pages1 s2.0 S0168169916303507 Main PDFa lNo ratings yet
- MitosisDocument7 pagesMitosisa lNo ratings yet
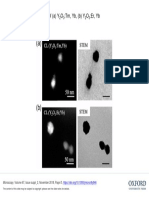
- Fig.1. CL and STEM Image of (A) Y: O:Tm, Yb, (B) Y O:Er, YbDocument1 pageFig.1. CL and STEM Image of (A) Y: O:Tm, Yb, (B) Y O:Er, Yba lNo ratings yet
- Everett Rogers Diffusion of Innovations Theory PDFDocument4 pagesEverett Rogers Diffusion of Innovations Theory PDFa lNo ratings yet
- TroubleshootingDocument9 pagesTroubleshootingNaja Maziyatul INo ratings yet
- 59-Article Text-396-1-10-20171124 PDFDocument8 pages59-Article Text-396-1-10-20171124 PDFSidik Taufik YusupNo ratings yet
- Experiment9 PlastidDocument4 pagesExperiment9 PlastidRahaizuddin RaniNo ratings yet
- Ultrastructure Nucleus: BiologyDocument14 pagesUltrastructure Nucleus: BiologyAshu ShewaleNo ratings yet
- Building Biotechnology 3rd EditionDocument24 pagesBuilding Biotechnology 3rd EditionPrincess Anne Lantin0% (1)
- Cell BingoDocument6 pagesCell BingoArcely DavidNo ratings yet
- Unit 7 - Viruses Bacteria Protists and FungiDocument63 pagesUnit 7 - Viruses Bacteria Protists and FungiGregson GucelaNo ratings yet
- Bacterial Classification and Morphology GuideDocument4 pagesBacterial Classification and Morphology GuideJerson Aizpuro SuplementoNo ratings yet
- Cytoplasmic InheritanceDocument11 pagesCytoplasmic InheritanceNaMan SeThiNo ratings yet
- Transplantation ImmunologyDocument18 pagesTransplantation Immunologyhendra2darmawanNo ratings yet
- MSc Biotechnology Scheme of Studies & Exams 2012-13Document39 pagesMSc Biotechnology Scheme of Studies & Exams 2012-13DeepakNo ratings yet
- Introduction To Cells PDFDocument6 pagesIntroduction To Cells PDFZainab HamidNo ratings yet
- LYSOSOMESDocument3 pagesLYSOSOMESAbdullah AbdullahNo ratings yet
- Edexcel Biology GCSE Topic 2 NotesDocument10 pagesEdexcel Biology GCSE Topic 2 NotesAbir OmarNo ratings yet
- Lakhmir Singh Biology Class 9 Solutions CH 2Document7 pagesLakhmir Singh Biology Class 9 Solutions CH 2DarshilNo ratings yet
- Vaidic Theory of Evolution Eng.Document9 pagesVaidic Theory of Evolution Eng.Atharv Arya100% (1)
- Cymat AluminiumDocument10 pagesCymat AluminiumnguyenvannganpcNo ratings yet
- BIO213 Fall 2016 SyllabusDocument4 pagesBIO213 Fall 2016 SyllabusJosephParkNo ratings yet
- Perfect Steps Publishers: End of Year Exams 2015Document7 pagesPerfect Steps Publishers: End of Year Exams 2015Jesse williamsNo ratings yet
- LET-PROF-ED-Prof ExerciseDocument15 pagesLET-PROF-ED-Prof ExerciseArmely Niedo100% (1)
- A General Biology I Q1M1.2Document26 pagesA General Biology I Q1M1.2Queng Eledia0% (1)
- Understanding Cell Structure and FunctionDocument2 pagesUnderstanding Cell Structure and FunctionNhs NhsNo ratings yet
- IAS - Biology - SB - Sample Topic 1 PDFDocument97 pagesIAS - Biology - SB - Sample Topic 1 PDFTed80% (5)
- Biological Sciences Drill Answer KeyDocument45 pagesBiological Sciences Drill Answer KeyanthonydongonNo ratings yet
- General Biology I M2Document26 pagesGeneral Biology I M2caiNo ratings yet
- EI 6704 Biomedical Instrumentation Unit IDocument69 pagesEI 6704 Biomedical Instrumentation Unit IInfant Raj100% (2)
- HIV (Human Immunodeficiency Virus) : 1. Binding and Fusion (Attachment)Document5 pagesHIV (Human Immunodeficiency Virus) : 1. Binding and Fusion (Attachment)mon chériNo ratings yet
- Virology: Structure and ReplicationDocument35 pagesVirology: Structure and ReplicationQuezon, Michaella BSN1No ratings yet
- Dwnload Full Psychology 8th Edition Gleitman Test Bank PDFDocument35 pagesDwnload Full Psychology 8th Edition Gleitman Test Bank PDFiseasfoqarae100% (10)
- Must-Know Key Concepts in PSLE ScienceDocument30 pagesMust-Know Key Concepts in PSLE ScienceClarisa CynthiaNo ratings yet
- Brock Microbiology 13 Lecture PresentationDocument80 pagesBrock Microbiology 13 Lecture PresentationFatma SagirNo ratings yet
- Dr. Retno Sutomo, PH.D, Sp.ADocument37 pagesDr. Retno Sutomo, PH.D, Sp.ADoni RevaiNo ratings yet