You might also like
- 2000 - Identification of Novel Human Genes Evolutionarily Conserved in Caenorhabditis Elegans by Comparative ProteomicsDocument11 pages2000 - Identification of Novel Human Genes Evolutionarily Conserved in Caenorhabditis Elegans by Comparative ProteomicsRaymond LaBoyNo ratings yet
- Mol Biol Evol 2011 Paar 1877 92Document16 pagesMol Biol Evol 2011 Paar 1877 92gunjan1920No ratings yet
- GenomicsDocument8 pagesGenomicsPRATAPNo ratings yet
- Fredrik Sterky and Joakim Lundeberg - Sequence Analysis of Genes and GenomesDocument31 pagesFredrik Sterky and Joakim Lundeberg - Sequence Analysis of Genes and GenomesYopghm698No ratings yet
- PR 50b7cd3dDocument21 pagesPR 50b7cd3dBinderiya EnkhbatNo ratings yet
- Analysis of The Human Tissue-Specific Expression by Genome-Wide Integration of Transcriptomics and Antibody-Based ProteomicsDocument10 pagesAnalysis of The Human Tissue-Specific Expression by Genome-Wide Integration of Transcriptomics and Antibody-Based ProteomicsJhonatan Stiven Ortega RamirezNo ratings yet
- Lincoln Stein - Genome Annotation: From Sequence To BiologyDocument13 pagesLincoln Stein - Genome Annotation: From Sequence To BiologyYopghm698No ratings yet
- Evolutionary Parameters of The Transcribed Mammalian GenomeDocument6 pagesEvolutionary Parameters of The Transcribed Mammalian GenomeLau Mandy KMNo ratings yet
- Genomics and Its Impact On Science and Society: A PrimerDocument12 pagesGenomics and Its Impact On Science and Society: A PrimerAdan VegaNo ratings yet
- Chapter 21 OutlineDocument21 pagesChapter 21 Outlineapi-270482617No ratings yet
- 2005 Antonell GenomeResDocument11 pages2005 Antonell GenomeResEAPNo ratings yet
- 12LectF Transcription 2017Document44 pages12LectF Transcription 2017Nofa RanaNo ratings yet
- Comparative GenomicsDocument1 pageComparative GenomicsSHRUTI SAMALNo ratings yet
- Tempo and Mode of Genome PDFDocument30 pagesTempo and Mode of Genome PDFGinna SolarteNo ratings yet
- Luận vănDocument23 pagesLuận vănHolmes SherlockNo ratings yet
- 1 s2.0 S0959437X1400094X MainDocument7 pages1 s2.0 S0959437X1400094X Maintemiska amatmuNo ratings yet
- 3 Model GenomesDocument4 pages3 Model GenomesParisha SinghNo ratings yet
- Identification and Characterization of Essential Genes in The Human GenomeDocument7 pagesIdentification and Characterization of Essential Genes in The Human Genome戴义宾No ratings yet
- Zhang 2012 The Evolution of Intron Size in Amniotes A Role For Powered Flight Genome Biol EvolDocument11 pagesZhang 2012 The Evolution of Intron Size in Amniotes A Role For Powered Flight Genome Biol EvolLaiaNo ratings yet
- Gorilla Genome Structural Variation Reveals Evolutionary Parallelisms With ChimpanzeeDocument10 pagesGorilla Genome Structural Variation Reveals Evolutionary Parallelisms With ChimpanzeeNiga LucianNo ratings yet
- Research Papers On Molecular Genetics PDFDocument7 pagesResearch Papers On Molecular Genetics PDFafmclccre100% (1)
- Human Genome ProjectDocument17 pagesHuman Genome Projectanon_306048523No ratings yet
- NIH Public Access: Author ManuscriptDocument9 pagesNIH Public Access: Author ManuscriptAndressa AlvesNo ratings yet
- Comp Transcript Between Zebrafsh and MammalsDocument5 pagesComp Transcript Between Zebrafsh and MammalsRita RodriguesNo ratings yet
- Adaptive Evolution of Foundation Kinetochore Proteins in PrimatesDocument13 pagesAdaptive Evolution of Foundation Kinetochore Proteins in Primates10sgNo ratings yet
- Proyecto Genoma HumanoDocument50 pagesProyecto Genoma HumanoLila Lubianka Domínguez RamírezNo ratings yet
- 12 BioDocument2 pages12 BioJdjdjxjdjNo ratings yet
- Black & White Deliver Card (1) - WPS OfficeDocument28 pagesBlack & White Deliver Card (1) - WPS Officelinkgogo69No ratings yet
- The Landscape of Human STR VariationDocument12 pagesThe Landscape of Human STR VariationJuliana SiruffoNo ratings yet
- Genomics and BioinformaticsDocument34 pagesGenomics and BioinformaticsEbook DownloadNo ratings yet
- Bio in For Ma TicsDocument8 pagesBio in For Ma TicsAdrian IvanNo ratings yet
- 6456456accepted Manuscript (Woerner)Document47 pages6456456accepted Manuscript (Woerner)RodrigoNo ratings yet
- Human GenomeDocument8 pagesHuman GenomeKannanSubramanianNo ratings yet
- Zebrafish and Other Non-Mammalian Models - Cristina PujadesDocument71 pagesZebrafish and Other Non-Mammalian Models - Cristina Pujadesmafe.z.solarte87No ratings yet
- Farm Animal Genomics and Informatics: An Update: Ahmed Fadiel, Ifeanyi Anidi and Kenneth D. EichenbaumDocument11 pagesFarm Animal Genomics and Informatics: An Update: Ahmed Fadiel, Ifeanyi Anidi and Kenneth D. EichenbaumFabian HdezNo ratings yet
- In Brief: Complexity On The NanoscaleDocument1 pageIn Brief: Complexity On The Nanoscaletopgun625No ratings yet
- Bioinformatics Class NotesDocument12 pagesBioinformatics Class NotesrajiNo ratings yet
- Transpo S OnesDocument8 pagesTranspo S Oneslalo199No ratings yet
- 10.1007@s11427 020 1709 6Document4 pages10.1007@s11427 020 1709 6Nouri RaidNo ratings yet
- T H G P: HE Uman Enome RojectDocument28 pagesT H G P: HE Uman Enome RojectUpendra Sharma Sulibele100% (1)
- Genome Res.-2018-Lee-1577-88Document13 pagesGenome Res.-2018-Lee-1577-88robbishNo ratings yet
- Is Junk DNA Bunk? A Critique of ENCODE: PerspectiveDocument7 pagesIs Junk DNA Bunk? A Critique of ENCODE: PerspectivemilithebillyNo ratings yet
- Biological Identifications Through DNA Barcodes - Cations Through DNA BarcodesDocument10 pagesBiological Identifications Through DNA Barcodes - Cations Through DNA BarcodesAndré Ampuero LeónNo ratings yet
- Lecture 2 TranscriptsDocument6 pagesLecture 2 TranscriptskittyngameNo ratings yet
- Initial Impact of The Sequencing of The Human GenomeDocument11 pagesInitial Impact of The Sequencing of The Human GenomeMariano PerezNo ratings yet
- Wiley American Journal of Botany: This Content Downloaded From 181.231.29.59 On Tue, 26 Mar 2019 21:10:07 UTCDocument17 pagesWiley American Journal of Botany: This Content Downloaded From 181.231.29.59 On Tue, 26 Mar 2019 21:10:07 UTCFranco SantinNo ratings yet
- 2021 12 08 471837 FullDocument39 pages2021 12 08 471837 FulljojojazzprogNo ratings yet
- Molecular Ecology - 2012 - O Neill - Parallel Tagged Amplicon Sequencing Reveals Major Lineages and Phylogenetic StructureDocument19 pagesMolecular Ecology - 2012 - O Neill - Parallel Tagged Amplicon Sequencing Reveals Major Lineages and Phylogenetic Structuremendozar787No ratings yet
- Oral Microbiology and Genomics: Periodontology 2000Document9 pagesOral Microbiology and Genomics: Periodontology 2000ammarstein89No ratings yet
- Pub - Encyclopedia of Genetics Genomics Proteomics and B PDFDocument4,046 pagesPub - Encyclopedia of Genetics Genomics Proteomics and B PDFraj252000100% (1)
- Article GenomicaDocument15 pagesArticle GenomicaLaiaNo ratings yet
- 2017 - Cre-loxP-Mediated Recombination - General Principles and Experimental ConsiderationsDocument12 pages2017 - Cre-loxP-Mediated Recombination - General Principles and Experimental ConsiderationsLucas PaulaNo ratings yet
- Mouse Genetic PortraitDocument24 pagesMouse Genetic PortraitbelovedaffectionNo ratings yet
- A New and Exciting Era of Genomics: No Region Is Beyond ReachDocument2 pagesA New and Exciting Era of Genomics: No Region Is Beyond ReachHaseena SaitNo ratings yet
- ATAC-seq A Method For Assaying Chromatin AccessibilityDocument10 pagesATAC-seq A Method For Assaying Chromatin AccessibilityWarriors96No ratings yet
- GenomeDocument26 pagesGenomeAldave MoonNo ratings yet
- CNV and Human HealthDocument4 pagesCNV and Human HealthThaina MattosNo ratings yet
- Principles of Transcriptome Analysis and Gene Expression Quantification: An RNA-seq TutorialDocument14 pagesPrinciples of Transcriptome Analysis and Gene Expression Quantification: An RNA-seq TutorialSujitha maryNo ratings yet
- Assignment 1 ProteomicsDocument3 pagesAssignment 1 ProteomicsSharifah NurainNo ratings yet
- Human Genome Structure, Function and Clinical ConsiderationsFrom EverandHuman Genome Structure, Function and Clinical ConsiderationsLuciana Amaral HaddadNo ratings yet
- Dev 140962Document13 pagesDev 140962Yamile A Rodríguez RiascosNo ratings yet
- Yuyama 2019Document13 pagesYuyama 2019Yamile A Rodríguez RiascosNo ratings yet
- 10 1038@nrg 2017 21Document15 pages10 1038@nrg 2017 21Yamile A Rodríguez RiascosNo ratings yet
- Pgen 0020029Document8 pagesPgen 0020029Yamile A Rodríguez RiascosNo ratings yet
- 2021 02 25 432844v1 FullDocument60 pages2021 02 25 432844v1 FullYamile A Rodríguez RiascosNo ratings yet
- Nova HaywardDocument18 pagesNova HaywardYamile A Rodríguez RiascosNo ratings yet
- 10 1016@j Cell 2016 03 034Document15 pages10 1016@j Cell 2016 03 034Yamile A Rodríguez RiascosNo ratings yet
- 1 s2.0 S0012160619304130 MainDocument12 pages1 s2.0 S0012160619304130 MainYamile A Rodríguez RiascosNo ratings yet
- Comparative Genomics and Phylogenomic Analysis of The Genus SalinivibrioDocument15 pagesComparative Genomics and Phylogenomic Analysis of The Genus SalinivibrioYamile A Rodríguez RiascosNo ratings yet
- NIH Public Access: Author ManuscriptDocument20 pagesNIH Public Access: Author ManuscriptYamile A Rodríguez RiascosNo ratings yet
- Dong-Chen2013 Article TranscriptomicsAdvancesAndApprDocument8 pagesDong-Chen2013 Article TranscriptomicsAdvancesAndApprYamile A Rodríguez RiascosNo ratings yet
- Nih Public Access: Map'Ing Cns Development and Cognition: An Erksome ProcessDocument15 pagesNih Public Access: Map'Ing Cns Development and Cognition: An Erksome ProcessYamile A Rodríguez RiascosNo ratings yet
- Genome Res.-2013-Alföldi-1063-8mmDocument11 pagesGenome Res.-2013-Alföldi-1063-8mmYamile A Rodríguez RiascosNo ratings yet
- Functional Enrichment Analysis Based On Long Noncoding RNA AssociationsDocument10 pagesFunctional Enrichment Analysis Based On Long Noncoding RNA AssociationsYamile A Rodríguez RiascosNo ratings yet
- Biological OrganizationDocument70 pagesBiological OrganizationSabrina LavegaNo ratings yet
- Home About Enthusiastic Endorsements Products & Services Soil Test Blooming BuddiesDocument10 pagesHome About Enthusiastic Endorsements Products & Services Soil Test Blooming BuddiesJelaiNo ratings yet
- Genetics of Endocrinology: Amit R. Majithia - David Altshuler - Joel N. HirschhornDocument20 pagesGenetics of Endocrinology: Amit R. Majithia - David Altshuler - Joel N. HirschhornDr Mehul Kumar ChourasiaNo ratings yet
- Food Storage in PlantsDocument5 pagesFood Storage in PlantsKeelah BennNo ratings yet
- Niko Tinbergen and The Red Patch On The Herring Gull's BeakDocument10 pagesNiko Tinbergen and The Red Patch On The Herring Gull's BeakchurianaNo ratings yet
- Biology Test - Grade 8 - Photosynthesis & Flowering Plants PDFDocument3 pagesBiology Test - Grade 8 - Photosynthesis & Flowering Plants PDFJaimeen GujarNo ratings yet
- Phylum ChordataDocument13 pagesPhylum ChordataEJ RamosNo ratings yet
- Reptiles Amphibians and Birds Tema 5Document1 pageReptiles Amphibians and Birds Tema 5pepeNo ratings yet
- The Trees of Florida - A Reference and Field Guide - Gil Nelson (1994) PDFDocument296 pagesThe Trees of Florida - A Reference and Field Guide - Gil Nelson (1994) PDFTruco El Martinez100% (1)
- Wild Animal Riddles Key Fun Activities Games Games Icebreakers Oneonone Ac - 118126Document2 pagesWild Animal Riddles Key Fun Activities Games Games Icebreakers Oneonone Ac - 118126Any BoraNo ratings yet
- Avaliação de Inglês 4° Ano - 4 - Bim - 2022Document3 pagesAvaliação de Inglês 4° Ano - 4 - Bim - 2022leila cristiane de limaNo ratings yet
- Science, Level 4, Practice 2Document5 pagesScience, Level 4, Practice 2vicky chanNo ratings yet
- BIO - F5 - KSSM - CHAPTER - 1 - 1.1 and 1.2 ORGANISATION OF PLANT TISSUES AND GROWTHDocument59 pagesBIO - F5 - KSSM - CHAPTER - 1 - 1.1 and 1.2 ORGANISATION OF PLANT TISSUES AND GROWTHwienna1987100% (2)
- Types of Plant DiseasesDocument14 pagesTypes of Plant DiseasesMuqadas NoorNo ratings yet
- Science 5 QTR 1Document4 pagesScience 5 QTR 1ISABELITANo ratings yet
- Evolution of Biotechnology Role of Chemical Engineer PDFDocument2 pagesEvolution of Biotechnology Role of Chemical Engineer PDFBarbNo ratings yet
- Bakker1992B BLUMEA1992 37 PDFDocument31 pagesBakker1992B BLUMEA1992 37 PDFJohn Marlon MagbuoNo ratings yet
- Canopy ManagementDocument19 pagesCanopy ManagementAtul RajNo ratings yet
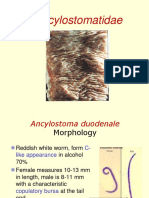
- Ancylostomatidae, T.trichura, S.stercoralisDocument19 pagesAncylostomatidae, T.trichura, S.stercoralisIqrinawidyazaharaNo ratings yet
- Jean Baptiste Lamarck and OthersDocument23 pagesJean Baptiste Lamarck and OthersShelley Mae YabesNo ratings yet
- History: The Value of Natural CollectionsDocument7 pagesHistory: The Value of Natural CollectionsMaruf PattimuraNo ratings yet
- Introduction To The Concept of Stem CellsDocument37 pagesIntroduction To The Concept of Stem Cellsscort cherNo ratings yet
- SummaryofreportmicrobioDocument5 pagesSummaryofreportmicrobiomysterioushumaneNo ratings yet
- Myrteae Phylogeny Calibration Biogeography and Div PDFDocument48 pagesMyrteae Phylogeny Calibration Biogeography and Div PDFJosé Caro LagosNo ratings yet
- Esr June 2019Document46 pagesEsr June 2019Gerd-Jan ten BrinkNo ratings yet
- Cambridge IGCSE (9-1) : Biology For Examination From 2023Document12 pagesCambridge IGCSE (9-1) : Biology For Examination From 2023valentinaNo ratings yet
- 1.biosphere Ecosystem and Biodiversity Loss PDFDocument221 pages1.biosphere Ecosystem and Biodiversity Loss PDFsalsabila100% (1)
- AmphibiansDocument17 pagesAmphibiansBONAVENTURE DEDENo ratings yet
- Biosphere Reserve 99Document6 pagesBiosphere Reserve 99Raman singh ChauhanNo ratings yet
- Ecology Demography and EpidemiologyDocument20 pagesEcology Demography and EpidemiologyNNNo ratings yet