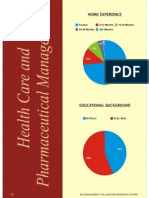
You might also like
- 1 s2.0 S0026049515003650 MainDocument12 pages1 s2.0 S0026049515003650 MainmmsNo ratings yet
- Clinical Nutrition ESPEN: Hiroshige Itakura, Michie Kobayashi, Seiji NakamuraDocument7 pagesClinical Nutrition ESPEN: Hiroshige Itakura, Michie Kobayashi, Seiji NakamuraPáblo EugênioNo ratings yet
- Cloves 1Document9 pagesCloves 1irmaNo ratings yet
- Jci37385 PDFDocument13 pagesJci37385 PDFNur Ahmad HabibiNo ratings yet
- Waterman Et Al-2015-Molecular Nutrition & Food ResearchDocument12 pagesWaterman Et Al-2015-Molecular Nutrition & Food ResearchIqra RubabNo ratings yet
- A Low-Fat Diet Decreases High Density Lipoprotein (HDL) Cholesterol Levels by Decreasing HDL Apolipoprotein Transport Rates. Brinton 1990Document8 pagesA Low-Fat Diet Decreases High Density Lipoprotein (HDL) Cholesterol Levels by Decreasing HDL Apolipoprotein Transport Rates. Brinton 1990acolpoNo ratings yet
- Background: Study DetailsDocument1 pageBackground: Study DetailsjoycechicagoNo ratings yet
- Am J Clin Nutr 2014 Lecoultre 268 75Document8 pagesAm J Clin Nutr 2014 Lecoultre 268 75Kang Opik TaufikNo ratings yet
- Ketogenic Diets Enhance Oxidative Stress and RadioChemo-Therapy Responses in Lung Cancer XenograftsDocument9 pagesKetogenic Diets Enhance Oxidative Stress and RadioChemo-Therapy Responses in Lung Cancer XenograftsCata AjuaNo ratings yet
- Asemi 2015Document7 pagesAsemi 2015ahmad azhar marzuqiNo ratings yet
- Peanut Consumption and Cardiovascular RiskDocument6 pagesPeanut Consumption and Cardiovascular RiskWahyuningsih HamidNo ratings yet
- Pomegranate Juice HealingDocument23 pagesPomegranate Juice HealingKingNo ratings yet
- Sunday O. Oyedemi - Godswill Nwaogu - Chika I. Chukwuma - Olaoluwa T. Adeyemi - Motlalepula G. Matsabisa - Shasank S. Swain - Olayinka A. AiyegoroDocument16 pagesSunday O. Oyedemi - Godswill Nwaogu - Chika I. Chukwuma - Olaoluwa T. Adeyemi - Motlalepula G. Matsabisa - Shasank S. Swain - Olayinka A. AiyegoroGiovanni OliveiraNo ratings yet
- Nutrients: Capsaicin Supplementation Improved Risk Factors of Coronary Heart Disease in Individuals With Low HDL-C LevelsDocument13 pagesNutrients: Capsaicin Supplementation Improved Risk Factors of Coronary Heart Disease in Individuals With Low HDL-C LevelsEmira LekaNo ratings yet
- BIOS LIFE - Cleveland Clinic Trial by Dr. Dennis SprecherDocument5 pagesBIOS LIFE - Cleveland Clinic Trial by Dr. Dennis SprecherHisWellnessNo ratings yet
- Art 3A10.1186 2Fs40795 017 0152 4Document9 pagesArt 3A10.1186 2Fs40795 017 0152 4NurfadhilahNo ratings yet
- Graessler 2012Document9 pagesGraessler 2012dora guzmanNo ratings yet
- 1826 Full PDFDocument7 pages1826 Full PDFNia Tiffany HarrkaNo ratings yet
- ComparisonDocument7 pagesComparisonJaqueline Driemeyer C. HorvathNo ratings yet
- Antiglycation and Hypolipidemic Effects of Polyphenols From Zingiber Officinale Roscoe (Zingiberaceae) in Streptozotocin-Induced Diabetic RatsDocument7 pagesAntiglycation and Hypolipidemic Effects of Polyphenols From Zingiber Officinale Roscoe (Zingiberaceae) in Streptozotocin-Induced Diabetic RatsJeli Jati AnggeriaNo ratings yet
- Clinical Trial 2016Document11 pagesClinical Trial 2016guugle gogleNo ratings yet
- Onion Peel Extracts Ameliorate Hyperglycemia and Insulin Resistance in High Fat Diet/ Streptozotocin-Induced Diabetic RatsDocument8 pagesOnion Peel Extracts Ameliorate Hyperglycemia and Insulin Resistance in High Fat Diet/ Streptozotocin-Induced Diabetic RatsRizky Agustian HadiNo ratings yet
- Nutrients 12 01781Document13 pagesNutrients 12 01781Barbara DumasNo ratings yet
- Regulation of Lipogenesis by Glucocorticoids and Insulin in Human Adipose TissueDocument9 pagesRegulation of Lipogenesis by Glucocorticoids and Insulin in Human Adipose TissuemamithemadNo ratings yet
- DiabetesDocument5 pagesDiabetespinaslifeNo ratings yet
- Shulman 2011Document8 pagesShulman 2011Alyna AlynaNo ratings yet
- Sugary Kefir Strain Lactobacillus Mali APS1 Ameliorated Hepatic Steatosis by Regulation of SIRT-1 NR-2 and Gut Microbiota in RatsDocument24 pagesSugary Kefir Strain Lactobacillus Mali APS1 Ameliorated Hepatic Steatosis by Regulation of SIRT-1 NR-2 and Gut Microbiota in RatsCristina OliveiraNo ratings yet
- A novel oligosaccharide isolated from Hericium erinaceus and its protection against LPS-induced Caco-2 cells via the TLR4 - NF-κB pathwayDocument9 pagesA novel oligosaccharide isolated from Hericium erinaceus and its protection against LPS-induced Caco-2 cells via the TLR4 - NF-κB pathwaymalvina.sekolonik911No ratings yet
- JN 212514Document6 pagesJN 212514Conciencia CristalinaNo ratings yet
- Nutrients 13 03965 v2Document11 pagesNutrients 13 03965 v2VaniNo ratings yet
- 2015 Article 45Document9 pages2015 Article 45Dr. Kaushal Kishor SharmaNo ratings yet
- Geetha 2013Document12 pagesGeetha 2013МарияNo ratings yet
- Journal About OatDocument8 pagesJournal About OatHegar JoNo ratings yet
- Effect of Wakame On Non Alcoholic Fatty LiverDocument14 pagesEffect of Wakame On Non Alcoholic Fatty LiverCătălina RădulescuNo ratings yet
- 879 FullDocument7 pages879 FullAhsan Tanio DaulayNo ratings yet
- Human Nutrition and MetabolismDocument6 pagesHuman Nutrition and MetabolismBruno CavicchioNo ratings yet
- Liraglutide Improves Lipid Metabolism by Enhancing Cholesterol Efflux Associated With ABCA1 and ERK1/2 PathwayDocument12 pagesLiraglutide Improves Lipid Metabolism by Enhancing Cholesterol Efflux Associated With ABCA1 and ERK1/2 PathwayAndrianna NastasyaNo ratings yet
- Bavane Et AlDocument5 pagesBavane Et AleditorijmrhsNo ratings yet
- Estudio Controlado de Fideos Instantaneos y Avena InstantaneaDocument8 pagesEstudio Controlado de Fideos Instantaneos y Avena InstantaneaChristian Cornejo RiveraNo ratings yet
- Journal of Separation Science: Version of Record Doi: 10.1002/jssc.202000911Document33 pagesJournal of Separation Science: Version of Record Doi: 10.1002/jssc.202000911Fabro BianNo ratings yet
- Human Nutrition and MetabolismDocument7 pagesHuman Nutrition and MetabolismMisyani MisyaniNo ratings yet
- Fatty FishDocument12 pagesFatty FishThục Ái Nguyễn NguyênNo ratings yet
- Am J Clin Nutr-2016-Samavat-1671-82Document12 pagesAm J Clin Nutr-2016-Samavat-1671-82ariesulaemanNo ratings yet
- AGES and NutritionDocument6 pagesAGES and NutritionBárbara Paranhos CoelhoNo ratings yet
- Effects of High Performance Inulin Supplementation On Glycemic Control and Antioxidant Status in Women With Type 2 DiabetesDocument9 pagesEffects of High Performance Inulin Supplementation On Glycemic Control and Antioxidant Status in Women With Type 2 DiabetesNia Tiffany HarrkaNo ratings yet
- Accepted Manuscript: 10.1016/j.metabol.2018.02.006Document26 pagesAccepted Manuscript: 10.1016/j.metabol.2018.02.006Rizqan Fahlevvi AkbarNo ratings yet
- Delgado Et Al 2009Document8 pagesDelgado Et Al 2009tcardosodelgadoNo ratings yet
- Biomedicine & Pharmacotherapy: SciencedirectDocument7 pagesBiomedicine & Pharmacotherapy: SciencedirectMirja AdiyaksaNo ratings yet
- L Argina PDFDocument6 pagesL Argina PDFAnyi VargasNo ratings yet
- Omega-3 Fatty Acids, Polymorphisms and Lipid Related Cardiovascular Disease Risk Factors in The Inuit PopulationDocument9 pagesOmega-3 Fatty Acids, Polymorphisms and Lipid Related Cardiovascular Disease Risk Factors in The Inuit Populationyoga diputraNo ratings yet
- Artículo Analogo (BCTP)Document7 pagesArtículo Analogo (BCTP)mrintraNo ratings yet
- Postprandial Plasma Lipidome Responses To A High-Fat Meal Among Healthy WomenDocument10 pagesPostprandial Plasma Lipidome Responses To A High-Fat Meal Among Healthy WomenMark DenhamNo ratings yet
- Increased Potassium Intake From Fruit and Vegetabl PDFDocument9 pagesIncreased Potassium Intake From Fruit and Vegetabl PDFburhangunawanNo ratings yet
- KersenDocument13 pagesKersenJumasni AdnanNo ratings yet
- Effect of Melatonin and Chromium Picolinate Administration To Highcarbohydrate Dietfed Male Wistar Rats 1747 0862 1000245Document7 pagesEffect of Melatonin and Chromium Picolinate Administration To Highcarbohydrate Dietfed Male Wistar Rats 1747 0862 1000245jovaneticNo ratings yet
- Accelerated Intestinal Glucose Absorption in Morbidly Obese Humans: Relationship To Glucose Transporters, Incretin Hormones, and GlycemiaDocument9 pagesAccelerated Intestinal Glucose Absorption in Morbidly Obese Humans: Relationship To Glucose Transporters, Incretin Hormones, and GlycemiaNissa SissariNo ratings yet
- The Antihyperlipidaemic and Hepatoprotective Effect of Ipomoea Batatas L. Leaves Extract in High-Fat Diet RatsDocument7 pagesThe Antihyperlipidaemic and Hepatoprotective Effect of Ipomoea Batatas L. Leaves Extract in High-Fat Diet RatsIJPHSNo ratings yet
- Research ArticleDocument12 pagesResearch ArticleKarthiNo ratings yet
- DMJ 37 465Document10 pagesDMJ 37 465sn4s7nyxcbNo ratings yet
- OPD and OT Days of Doctors of Clinical Department of Guru Gobind Singh Medical HospitalDocument4 pagesOPD and OT Days of Doctors of Clinical Department of Guru Gobind Singh Medical HospitalBohar singh Brar BrarNo ratings yet
- Insanity Workout DVDDocument2 pagesInsanity Workout DVDshopzillahouse001No ratings yet
- Ansi A10.42Document23 pagesAnsi A10.42pedro la rosa100% (1)
- Health ChemoDocument9 pagesHealth ChemoKanjarla Hanmonthrao Panthulu LaxmanaNo ratings yet
- Icd 10 Rajal Poli SarafDocument3 pagesIcd 10 Rajal Poli Sarafgrace efiliaNo ratings yet
- Temporomandibular Disorders: A Term Whose Time Has Passed!: PerspectivesDocument2 pagesTemporomandibular Disorders: A Term Whose Time Has Passed!: PerspectivesUmer HussainNo ratings yet
- Ficha Tecnica E803Document2 pagesFicha Tecnica E803Zenen AlarconNo ratings yet
- Organ DonationDocument4 pagesOrgan DonationKrishna TejavathNo ratings yet
- DRUG StudyDocument43 pagesDRUG StudyNathalie Faith CotengNo ratings yet
- Case:: Biddy's Bakery (BB)Document3 pagesCase:: Biddy's Bakery (BB)tehreem chohanNo ratings yet
- Literature Review On OgiDocument7 pagesLiterature Review On Ogiafdtaeczo100% (1)
- Gymnemic AcidDocument9 pagesGymnemic AcidarumpratiwiNo ratings yet
- Allianz Care Plus Brochure Update 19mar2015 FA R3 5Document11 pagesAllianz Care Plus Brochure Update 19mar2015 FA R3 5Leonard Yang0% (1)
- HIV Peer Training Toolkit GuideDocument73 pagesHIV Peer Training Toolkit GuideGatot SandyNo ratings yet
- DLL - Mapeh 5 - Q3 - W6Document7 pagesDLL - Mapeh 5 - Q3 - W6Reniel SabacoNo ratings yet
- IES College PlacementDocument10 pagesIES College PlacementpankajcpatelNo ratings yet
- Tangina Mo BizarDocument1 pageTangina Mo BizarJhon eric EscultorNo ratings yet
- 22 Ref Doc VH202 White Paper PART2 Draft05Document6 pages22 Ref Doc VH202 White Paper PART2 Draft05Gaelle BaudouinNo ratings yet
- Screw Air Compressor L07-L22: User ManualDocument56 pagesScrew Air Compressor L07-L22: User ManualЯни Хандрулев100% (4)
- Performance Evaluation HeadnursingDocument3 pagesPerformance Evaluation HeadnursingBing58No ratings yet
- POCKIT Central Nucleic Acid Analyzer (CE-IVD)Document2 pagesPOCKIT Central Nucleic Acid Analyzer (CE-IVD)Fadli AmbaraNo ratings yet
- Short Bowel SyndromeDocument20 pagesShort Bowel SyndromeMohamed El-sayedNo ratings yet
- 46TANASESCU918Document4 pages46TANASESCU918Fuja AmandaNo ratings yet
- City of Houston Storm Water QualityDocument45 pagesCity of Houston Storm Water Qualitysmcguire6631No ratings yet
- Turkish Exhibitors: 8-11 MARCH 2013Document9 pagesTurkish Exhibitors: 8-11 MARCH 2013qday1No ratings yet
- By Dinesh GannerllaDocument74 pagesBy Dinesh GannerllagannerllaNo ratings yet
- (09032021) Press Release For Resumption of Selected Sports For NSG 2021 - MOEDocument2 pages(09032021) Press Release For Resumption of Selected Sports For NSG 2021 - MOEthesuperprogamer2 gamerNo ratings yet
- Jerusalem SyndromeDocument5 pagesJerusalem Syndromele_papillon15No ratings yet
- DISABILITYDocument9 pagesDISABILITYSyeda AttqaNo ratings yet
- HPLC Analysis of AlprazolamDocument8 pagesHPLC Analysis of AlprazolamJuan PerezNo ratings yet